Citation: Viktorija Dičpinigaitienė, Lina Novickytė. Application of systemic risk measurement methods: A systematic review and meta-analysis using a network approach[J]. Quantitative Finance and Economics, 2018, 2(4): 798-820. doi: 10.3934/QFE.2018.4.798

| [1] |
Abdymomunov A (2013) Regime-switching measure of systemic financial stress. Ann Finance 9: 455–470. doi: 10.1007/s10436-012-0194-1

|
| [2] |
Acharya V, Engle R, Pierret D (2014) Testing macroprudential stress tests: The risk of regulatory risk weights. J Monetary Econ 65: 36–53. doi: 10.1016/j.jmoneco.2014.04.014
|
| [3] | Adrian T, Brunnermeier MK (2011) CoVaR. NBER, Working Paper, 17454. |
| [4] | Aikman D, Kiley M, Lee SJ, et al. (2017) Mapping heat in the U.S. financial system. J Banking Financ 81: 36–64. |
| [5] |
Aldasoro I, Angeloni I (2015) Input-output-based measures of systemic importance. Quant Financ 15: 589–606. doi: 10.1080/14697688.2014.968194
|
| [6] | Anginer D, Demirguc-Kunt A, Zhu M (2014) How does competition affect bank systemic risk? J Financ Int 23: 1–26. |
| [7] |
Baek S, Cursio JD, Cha SY (2015) Nonparametric Factor Analytic Risk Measurement in Common Stocks in Financial Firms: Evidence from Korean Firms. Asia-Pac J Financ Stud 44: 497–536. doi: 10.1111/ajfs.12098
|
| [8] |
Baglioni A, Cherubini U (2013) Marking-to-market government guarantees to financial systems-Theory and evidence for Europe. J Int Money and Financ 32: 990–1007. doi: 10.1016/j.jimonfin.2012.08.004
|
| [9] | Banulescu GD, Dumitrescu EI (2015) Which are the SIFIs? A Component Expected Shortfall approach to systemic risk. J Banking Financ 50: 575–588. |
| [10] | Barth JR, Wihlborg C (2017) Too big to fail: Measures, remedies, and consequences for efficiency and stability. Financ Mark Inst Instrum 26: 175–245. |
| [11] |
Battaglia F, Gallo A (2013) Securitization and systemic risk: An empirical investigation on Italian banks over the financial crisis. Int Rev Financ Anal 30: 274–286. doi: 10.1016/j.irfa.2013.03.002
|
| [12] | Battaglia F, Gallo A (2017) Strong boards, ownership concentration and EU banks' systemic risk-taking: Evidence from the financial crisis. J Int Financ Mark 46: 128–146. |
| [13] | Benoit S (2014) Where is the system? Int Econ 138: 1–27. |
| [14] | Berger D, Pukthuanthong K (2012) Market fragility and international market crashes. J Financ Econ 105: 565–580. |
| [15] | Bernal O, Gnabo JY, Guilmin G (2014) Assessing the contribution of banks, insurance and other financial services to systemic risk. J Banking Financ 47: 270–287. |
| [16] |
Bernardi M, Maruotti A, Petrella L (2017) Multiple risk measures for multivariate dynamic heavy-tailed models. J Empirical Financ 43: 1–32. doi: 10.1016/j.jempfin.2017.04.005
|
| [17] |
Bisias D, Flood M, Lo AW, et al. (2012) A survey of systemic risk analytics. Annu Rev Financ Econ 4: 255–296. doi: 10.1146/annurev-financial-110311-101754
|
| [18] |
Bluhm M, Krahnen JP (2014) Systemic risk in an interconnected banking system with endogenous asset markets. J Financ Stab 13: 75–94. doi: 10.1016/j.jfs.2014.04.002
|
| [19] | Borri N (2017) Local currency systemic risk. Emerging Mark Rev 34: 1–13. |
| [20] | Cai J, Eidam F, Saunders A, et al. (2018) Syndication, interconnectedness, and systemic risk. J Financ Stab 34: 105–120. |
| [21] |
Cambón MI, Estévez L (2016) A Spanish Financial Market Stress Index (FMSI). Span Rev Financ Econ 14: 23–41. doi: 10.1016/j.srfe.2016.01.002
|
| [22] | Castro C, Ferrari S (2014) Measuring and testing for the systemically important financial institutions. J Empirical Financ 25: 1–14. |
| [23] | Cerchiello P, Giudici P (2016) Big data analysis for financial risk management. J Big Data 3: 18. |
| [24] |
Cerchiello P, Giudici P, Nicola G (2017) Twitter data models for bank risk contagion. Neurocomputing 264: 50–56. doi: 10.1016/j.neucom.2016.10.101
|
| [25] | Chakroun MA, Gallali MI (2017) Contribution of Islamic banks to systemic risk. Int J Banking Accounting 8: 52–92. |
| [26] | Chang CW, Li X, Lin EMH, et al. (2017) Systemic risk, interconnectedness, and non-core activities in Taiwan insurance industry. Int Rev Econ Financ 55: 1–12. |
| [27] | Cheng F, Wellman MP (2017) Accounting for strategic response in an agent-based model of financial regulation. Acm Conf Econ Comput 2017: 187–204. |
| [28] | Chiu WC, Peña JI, Wang CW (2015) Measuring Systemic Risk: Common Factor Exposures and Tail Dependence Effects. Eur Financ Manage 21: 833–866. |
| [29] | Cipra T, Hendrych R (2017) Systemic risk in financial risk regulation. Czech J Econ Financ 67: 15–38. |
| [30] | Conciarelli A (2014) A New macroprudential tool to assess sources of financial risks: Implied-systemic cost of risks. Int J Financ Econ 19: 74–88. |
| [31] | Derbali A (2017) Systemic Risk in the Chinese Financial System: Measuring and Ranking. Chin Econ 50: 34–58. |
| [32] | Derbali A, Hallara S (2016) Systemic risk of European financial institutions: Estimation and ranking by the Marginal Expected Shortfall. Res Int Bus Financ 37: 113–134. |
| [33] | Dhar V (2013) Data science and prediction. Commun ACM 56: 64–73. |
| [34] | Cesare AD, Stork PA, De Vries CG (2012) Risk measures for autocorrelated hedge fund returns. J Financ Econ 13: 868–895. |
| [35] | Drakos AA, Kouretas GP (2013) Measuring Systemic Risk in Emerging Markets Using CoVaR. Emerging Markets and the Global Economy: A Handbook, 271–307. |
| [36] |
Drakos AA, Kouretas GP (2015) Bank ownership, financial segments and the measurement of systemic risk: An application of CoVaR. Int Rev Econ Financ 40: 127–140. doi: 10.1016/j.iref.2015.02.010
|
| [37] | Drehmann M, Tarashev N (2013) Measuring the systemic importance of interconnected banks. J Financ Int 22: 586–607. |
| [38] | Foggitt GM, Heymans A, Vuuren GWV, et al. (2017) Measuring the systemic risk in the South African banking sector. South Afr J Econ Manage Sci 20: 1–9. |
| [39] |
Garciadeandoain C, Kremer M (2017) Beyond spreads: Measuring sovereign market stress in the euro area. Econ Lett 159: 153–156. doi: 10.1016/j.econlet.2017.06.042
|
| [40] | Gauthier C, Lehar A, Souissi M (2012) Macroprudential capital requirements and systemic risk. J Financ Int 21: 594–618. |
| [41] |
Girardi G, Ergün AT (2013) Systemic risk measurement: Multivariate GARCH estimation of CoVaR. J Banking Financ 37: 3169–3180. doi: 10.1016/j.jbankfin.2013.02.027
|
| [42] |
Giudici P, Parisi L (2017) Sovereign risk in the Euro area: A multivariate stochastic process approach. Quant Financ 17: 1995–2008. doi: 10.1080/14697688.2017.1357968
|
| [43] | González-Hermosillo B, Hesse H (2011) Global Market Conditions and Systemic Risk. J Emerging Mark Financ 10: 227–252. |
| [44] | Gramlich D, Oet MV, Ong SJ (2017) The contributions to systemic stress of financial interactions between the US and Europe. Eur J Financ 23: 1176–1196. |
| [45] | Gray DF, Malone SW (2012) Sovereign and financial-sector risk: Measurement and interactions. Annu Rev Financ Econ 4: 297–312. |
| [46] | Hałaj G, Kok C (2013) Assessing interbank contagion using simulated networks. Comput Manage Sci 10: 157–186. |
| [47] | Härdle WK, Wang W, Yu L (2016) TENET: Tail–event driven NETwork risk. J Econ 192: 499–513. |
| [48] |
Hespeler F, Loiacono G (2017) Monitoring systemic risk in the hedge fund sector. Quant Financ 17: 1859–1883. doi: 10.1080/14697688.2017.1357969
|
| [49] |
Hmissi B, Bejaoui A, Snoussi W (2017) On identifying the domestic systemically important banks: The case of Tunisia. Res Int Bus Financ 42: 1343–1354. doi: 10.1016/j.ribaf.2017.07.071
|
| [50] |
Huang X, Zhou H, Zhu H (2009) A framework for assessing the systemic risk of major financial institutions. J Banking Financ 33: 2036–2049. doi: 10.1016/j.jbankfin.2009.05.017
|
| [51] |
Huang X (2012) Systemic risk contributions. J Financ Serv Res 42: 55–83. doi: 10.1007/s10693-011-0117-8
|
| [52] | IJtsma P, Spierdijk L, Shaffer S (2017) The concentration-stability controversy in banking: New evidence from the EU-25. J Financ Stab 33: 273–284. |
| [53] | Irresberger F, Bierth C, Weiß GNF (2017) Size is everything: Explaining SIFI designations. Rev Financi Econ 32: 7–19. |
| [54] | Jobst AA (2014) Measuring systemic risk-adjusted liquidity (SRL)-a model approach. J Banking Financ 45: 270–287. |
| [55] |
Jondeau E, Khalilzadeh A (2017) Collateralization, leverage, and stressed expected loss. J Financ Stab 33: 226–243. doi: 10.1016/j.jfs.2017.01.005
|
| [56] | Kanno M (2015a) Assessing systemic risk using interbank exposures in the global banking system. J Financ Stab 20: 105–130. |
| [57] | Kanno M (2015b) The network structure and systemic risk in the Japanese interbank market. Jpn World Econ 36: 102–112. |
| [58] |
Kanno M (2016) The network structure and systemic risk in the global non-life insurance market. Insur Math Econ 67: 38–53. doi: 10.1016/j.insmatheco.2015.12.004
|
| [59] | Karimalis EN, Nomikos NK (2017) Measuring systemic risk in the European banking sector: A copula CoVaR approach. Eur J Financ 24: 1–38. |
| [60] | Khiari W, Nachnouchi J (2017) Banks' systemic risk in the Tunisian context: Measures and Determinants. Res Int Bus Financ. |
| [61] | Kleinow J, Horsch A, Garcia-Molina M (2017a) Factors driving systemic risk of banks in Latin America. J Econ Financ 41: 211–234. |
| [62] | Kleinow J, Moreira F, Strobl S, et al. (2017b) Measuring systemic risk: A comparison of alternative market-based approaches. Financ Res Lett 21: 40–46. |
| [63] | Klinger T, Teplý P (2014) Systemic risk of the global banking system-an agent-based network model approach. Prague Econ Pap 23: 24–41. |
| [64] | Klinger T, Teplý P (2016) The nexus between systemic risk and sovereign crises. Czech J Econ Financ 66: 50–69. |
| [65] | Kreis Y, Leisen DPJ (2016) Systemic risk in a structural model of bank default linkages. J Financ Stab, 1–46. |
| [66] | Kubinschi M, Barnea D (2016) Systemic risk impact on economic growth-The case of the CEE countries. Rom J Econ Forecast 19: 79–94. |
| [67] | Kupiec PH, Ramirez CD (2013) Bank failures and the cost of systemic risk: Evidence from 1900 to 1930. J Financ Int 22: 285–307. |
| [68] | Kupiec P, Güntay L (2016) Testing for Systemic Risk Using Stock Returns. J Financ Serv Res 49: 203–227. |
| [69] |
Kurowski ŁK, Rogowicz K (2017) Negative interest rates as systemic risk event. Financ Res Lett 22: 153–157. doi: 10.1016/j.frl.2017.04.001
|
| [70] |
Lee JH, Ryu J, Tsomocos D (2013) Measures of systemic risk and financial fragility in Korea. Ann Financ 9: 757–786. doi: 10.1007/s10436-012-0218-x
|
| [71] | Lee J, Lee DH, Yun SG (2016) Systemic Risk on Trade Credit Systems: With the Tangible Interconnectedness. Comput Econ 51: 1–16. |
| [72] |
Leroy A, Lucotte Y (2017) Is there a competition–stability trade–off in European banking? J Int Financ Mark 46: 199–215. doi: 10.1016/j.intfin.2016.08.009
|
| [73] | Li F, Perez-Saiz H (2018) Measuring systemic risk across financial market infrastructures. J Financ Stab 34: 1–11. |
| [74] |
Liao S, Sojli E, Tham WW (2015) Managing systemic risk in The Netherlands. Int Rev Econ Financ 40: 231–245. doi: 10.1016/j.iref.2015.02.012
|
| [75] |
López-Espinosa G, Moreno A, Rubia A, et al. (2015) Systemic risk and asymmetric responses in the financial industry. J Banking Financ 58: 471–485. doi: 10.1016/j.jbankfin.2015.05.004
|
| [76] |
López-Espinosa G, Rubia A, Valderrama L, et al. (2013) Good for one, bad for all: Determinants of individual versus systemic risk. J Financ Stab 9: 287–299. doi: 10.1016/j.jfs.2013.05.002
|
| [77] |
Ma Y, Chen Y (2014) Financial imbalance index as a new early warning indicator: Methods and applications in the Chinese economy. China World Econ 22: 64–86. doi: 10.1111/cwe.12092
|
| [78] | Martinez-Jaramillo S, Alexandrova-Kabadjova B, Bravo-Benitez B, et al. (2014) An empirical study of the Mexican banking system's network and its implications for systemic risk. J Econ Dyn Control 40: 242–265. |
| [79] | Martínez-Jaramillo S, Pérez OP, Embriz FA, et al. (2010) Systemic risk, financial contagion and financial fragility. J Econ Dyn Control 34: 2358–2374. |
| [80] |
Mayordomo S, Rodriguez-Moreno M, Peña JI (2014) Derivatives holdings and systemic risk in the U.S. banking sector. J Banking Financ 45: 84–104. doi: 10.1016/j.jbankfin.2014.03.037
|
| [81] |
Mendonça HF, Silva RBD (2018) Effect of banking and macroeconomic variables on systemic risk: An application of Δ COVAR for an emerging economy. North Am J Econ Financ 43: 141–157. doi: 10.1016/j.najef.2017.10.011
|
| [82] | Mensah JO, Premaratne G (2017) Systemic interconnectedness among Asian Banks. Jpn World Econ 41: 17–33. |
| [83] | Mezei J, Sarlin P (2016) Aggregating expert knowledge for the measurement of systemic risk. Decis Support Syst 88: 38–50. |
| [84] |
Mezei J, Sarlin P (2018) RiskRank: Measuring interconnected risk. Econ Modell 68: 41–50. doi: 10.1016/j.econmod.2017.04.016
|
| [85] | Milne A (2014) Distance to default and the financial crisis. J Financ Stab 12: 26–36. |
| [86] | Nucera F, Lucas A, et al. (2017) Do negative interest rates make banks less safe? Econ Lett 159: 112–115. |
| [87] |
Patro DK, Qi M, Sun X (2013) A simple indicator of systemic risk. J Financ Stab 9: 105–116. doi: 10.1016/j.jfs.2012.03.002
|
| [88] |
Pederzoli C, Torricelli C (2017) Systemic risk measures and macroprudential stress tests: An assessment over the 2014 EBA exercise. Ann Financ 13: 237–251. doi: 10.1007/s10436-017-0294-z
|
| [89] | Popescu A, Turcu C (2014) Systemic sovereign risk in Europe: An MES and CES approach. Work Pap 124: 899–925. |
| [90] |
Popescu A, Turcu C (2017) Sovereign debt and systemic risk in the eurozone: A macroeconomic perspective. Econ Modell 67: 275–284. doi: 10.1016/j.econmod.2016.12.032
|
| [91] |
Reboredo JC, Ugolini A (2015) Systemic risk in European sovereign debt markets: A CoVaR-copula approach. J Int Money Financ 51: 214–244. doi: 10.1016/j.jimonfin.2014.12.002
|
| [92] | Reboredo JC, Ugolini A (2016) Systemic risk of Spanish listed banks: A vine copula CoVaR approach [Riesgo sistémico de los bancos españoles cotizados: Una aproximación CoVaR con cópulas vine]. Span J Financ Accounting 45: 1–31. |
| [93] |
Rönnqvist S, Sarlin P (2015) Bank networks from text: Interrelations, centrality and determinants. Quant Financ 15: 1619–1635. doi: 10.1080/14697688.2015.1071076
|
| [94] | Rösch D, Scheule H (2016) The role of loan portfolio losses and bank capital for Asian financial system resilience. Pac-Basin Financ J 40: 289–305. |
| [95] |
Sedunov J (2016) What is the systemic risk exposure of financial institutions? J Financ Stab 24: 71–87. doi: 10.1016/j.jfs.2016.04.005
|
| [96] | Sheu HJ, Cheng CL (2012) Systemic risk in Taiwan stock market. J Bus Econ Manage 13: 895–914. |
| [97] |
Siebenbrunner C, Sigmund M, Kerbl S (2017) Can bank-specific variables predict contagion effects? Quant Financ 17: 1805–1832. doi: 10.1080/14697688.2017.1357974
|
| [98] |
Silva TC, Souza SRSD, Tabak BM (2017) Monitoring vulnerability and impact diffusion in financial networks. J Econ Dyn Control 76: 109–135. doi: 10.1016/j.jedc.2017.01.001
|
| [99] |
Souza SRSD, Silva TC, Tabak BM, et al. (2016) Evaluating systemic risk using bank default probabilities in financial networks. J Econ Dyn Control 66: 54–75. doi: 10.1016/j.jedc.2016.03.003
|
| [100] |
Staum J (2012) Systemic risk components and deposit insurance premia. Quant Financ 12: 651–662. doi: 10.1080/14697688.2012.664942
|
| [101] | Stolbov M, Shchepeleva M (2017) Systemic risk in Europe: Deciphering leading measures, common patterns and real effects. Ann Financ 14: 1–43. |
| [102] |
Strobl S (2016) Stand-alone vs systemic risk-taking of financial institutions. J Risk Financ 17: 374–389. doi: 10.1108/JRF-05-2016-0064
|
| [103] |
Trabelsi N, Naifar N (2017) Are Islamic stock indexes exposed to systemic risk? Multivariate GARCH estimation of CoVaR. Res Int Bus Financ 42: 727–744. doi: 10.1016/j.ribaf.2017.07.013
|
| [104] |
Xu S, In F, Forbes C, et al. (2017) Systemic risk in the European sovereign and banking system. Quant Financ 17: 633–656. doi: 10.1080/14697688.2016.1205212
|
| [105] |
Yao Y, Li J, Zhu X, et al. (2017) Expected default based score for identifying systemically important banks. Econ Modell 64: 589–600. doi: 10.1016/j.econmod.2017.04.023
|
| [106] |
Yun J, Moon H (2014) Measuring systemic risk in the Korean banking sector via dynamic conditional correlation models. Pac-Basin Financ J 27: 94–114. doi: 10.1016/j.pacfin.2014.02.005
|
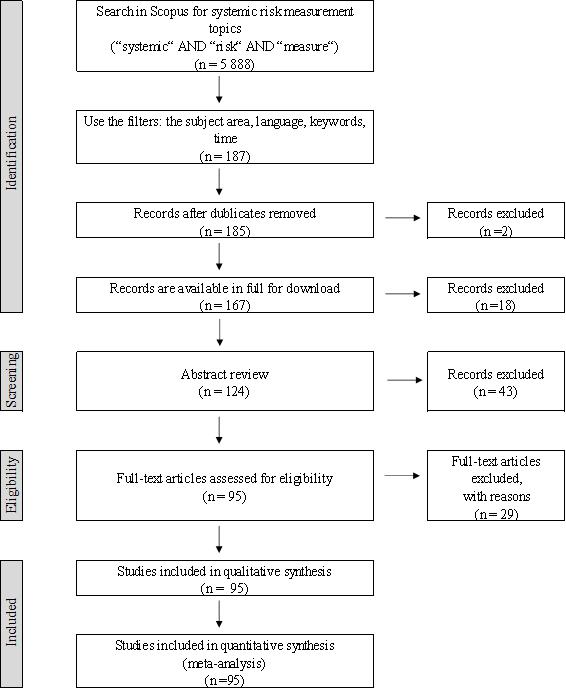
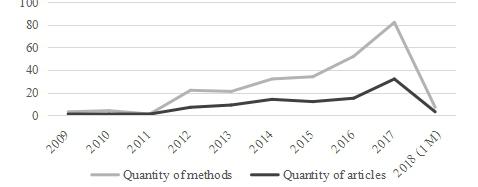
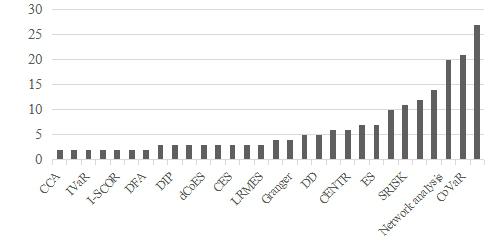
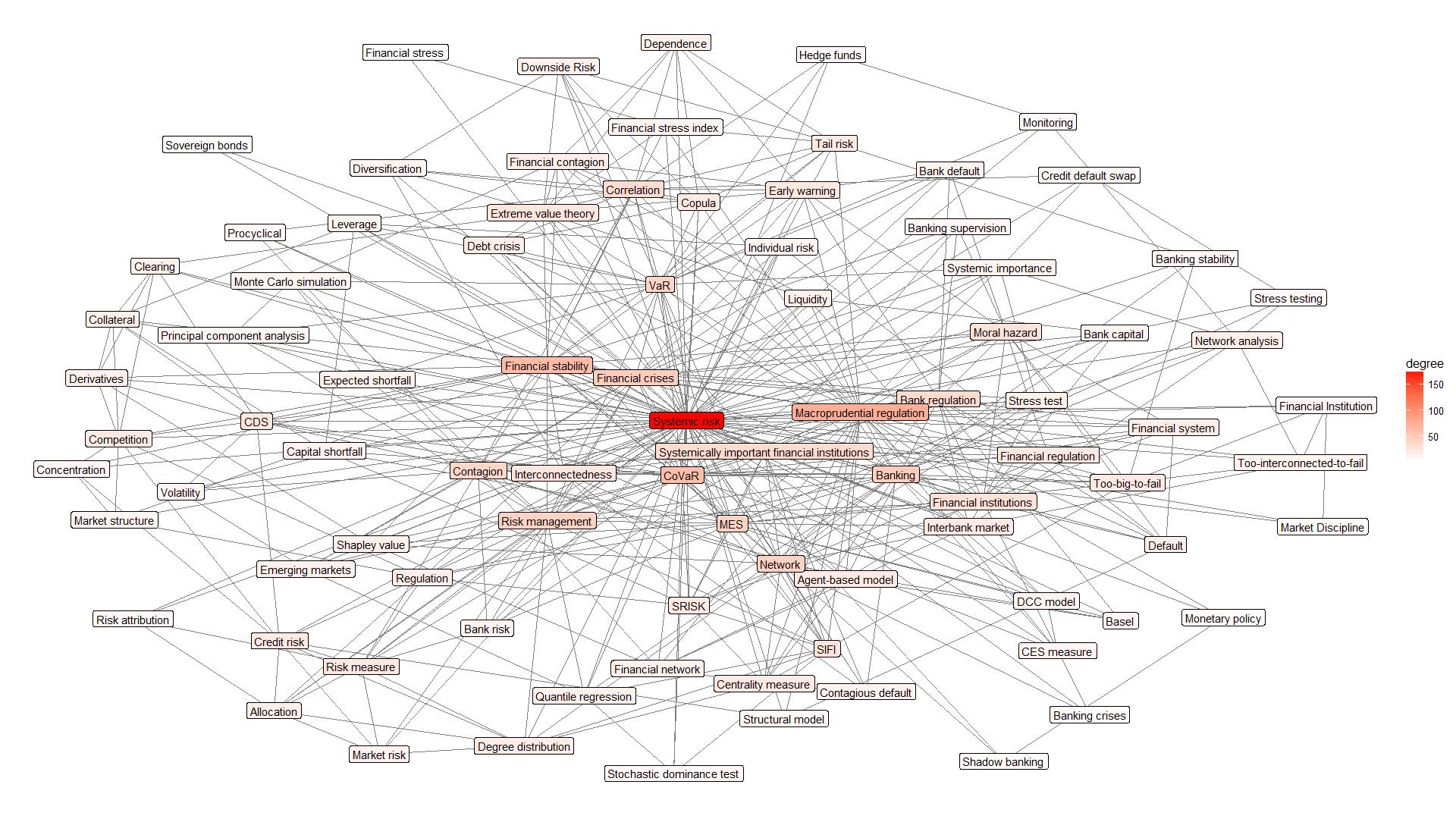

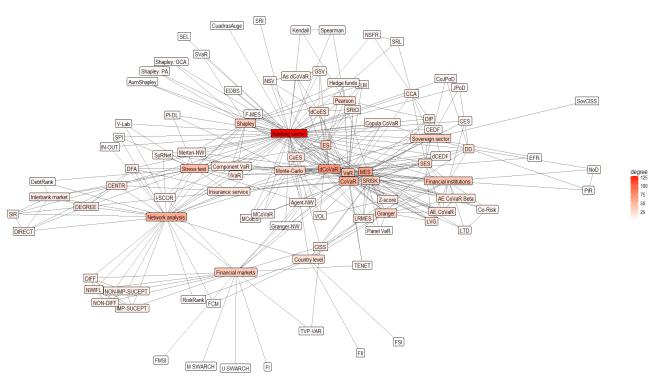
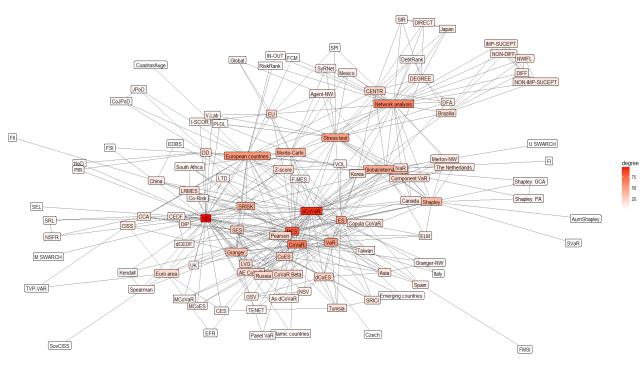
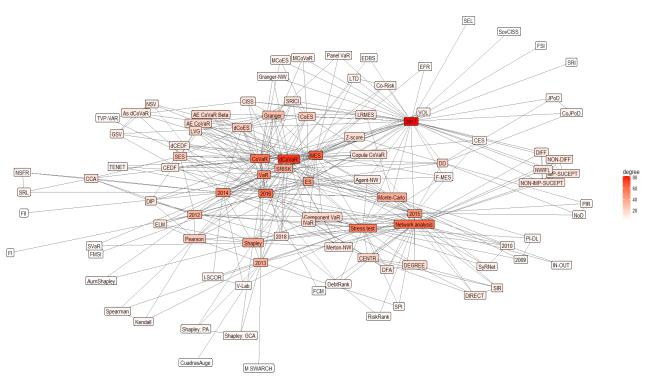
Figures(8) / Tables(2)

Viktorija Dičpinigaitienė, Lina Novickytė. Application of systemic risk measurement methods: A systematic review and meta-analysis using a network approach[J]. Quantitative Finance and Economics, 2018, 2(4): 798-820. doi: 10.3934/QFE.2018.4.798







 DownLoad:
DownLoad: