Citation: Eduardo Mercado III. Relating Cortical Wave Dynamics to Learning and Remembering[J]. AIMS Neuroscience, 2014, 1(3): 185-209. doi: 10.3934/Neuroscience.2014.3.185

| [1] | Takeuchi T, Duszkiewicz AJ, Morris RG (2014) The synaptic plasticity and memory hypothesis: Encoding, storage and persistence. Philos Trans R Soc Lond B Biol Sci 369: 20130288. |
| [2] |
Gallistel CR, Matzel LD (2013) The neuroscience of learning: Beyond the Hebbian synapse. Annu Rev Psychol 64: 169-200. doi: 10.1146/annurev-psych-113011-143807

|
| [3] |
Gallistel CR, Balsam PD (2014) Time to rethink the neural mechanisms of learning and memory. Neurobiol Learn Mem 108: 136-144. doi: 10.1016/j.nlm.2013.11.019
|
| [4] | Gallistel CR, King AP (2009) Memory and the computational brain: Why cognitive science will transform neuroscience. Chichester, UK: Wiley-Blackwell. |
| [5] |
Lynch G, Baudry M (1984) The biochemistry of memory: A new and specific hypothesis. Science 224: 1057-1063. doi: 10.1126/science.6144182
|
| [6] |
Baudry M, Bi X, Gall C, Lynch G (2011) The biochemistry of memory: The 26 year journey of a 'new and specific hypothesis'. Neurobiol Learn Mem 95: 125-133. doi: 10.1016/j.nlm.2010.11.015
|
| [7] |
Aimone JB, Deng W, Gage FH (2011) Resolving new memories: A critical look at the dentate gyrus, adult neurogenesis, and pattern separation. Neuron 70: 589-596. doi: 10.1016/j.neuron.2011.05.010
|
| [8] |
Buzsaki G (2010) Neural syntax: Cell assemblies, synapsembles, and readers. Neuron 68:362-385. doi: 10.1016/j.neuron.2010.09.023
|
| [9] | Pavlov IP (1927) Conditioned reflexes. London: Oxford University Press. |
| [10] |
Frostig RD, Xiong Y, Chen-Bee CH, et al. (2008) Large-scale organization of rat sensorimotor cortex based on a motif of large activation spreads. J Neurosci 28: 13274-13284. doi: 10.1523/JNEUROSCI.4074-08.2008
|
| [11] | Sechenov I (1965) Reflexes of the brain. Cambridge: MIT Press. |
| [12] | Sherrington CS (1906) The integrative action of the nervous system. New Haven: Yale University Press. |
| [13] | Jennings HS (1962) Behavior of the lower organisms. Bloomington: Indiana University Press. |
| [14] |
Spence KW (1937) The differential response in animals to stimuli varying within a single dimension. Psychol Rev 44: 430-444. doi: 10.1037/h0062885
|
| [15] |
Guthrie ER (1934) Discussion: Pavlov's theory of conditioning. Psychol Rev 41: 199-206. doi: 10.1037/h0071458
|
| [16] |
Wenger MA (1937) A criticism of Pavlov's concept of internal inhibition. Psychol Rev 44:297-312. doi: 10.1037/h0061974
|
| [17] |
Lashley KS, Wade M (1946) The Pavlovian theory of generalization. Psychol Rev 53: 72-87. doi: 10.1037/h0059999
|
| [18] | Denny-Brown D (1932) Critical review: Theoretical deductions from the physiology of the cerebral cortex. J Neurol Psychopathol 13: 52-67. |
| [19] |
Loucks RB (1937) Reflexology and the psychobiological approach. Psychol Rev 44: 320-338. doi: 10.1037/h0060693
|
| [20] |
Razran G (1949) Stimulus generalization of conditioned responses. Psychol Bull 46: 337-365. doi: 10.1037/h0060507
|
| [21] | Hebb DO (1949) The organization of behavior. New York: Wiley. |
| [22] | James W (1890) The principles of psychology. New York: Dover. |
| [23] |
24. Hughes JR, Ikram A, Fino JJ (1995). Characteristics of travelling waves under various conditions. Clin Electroencephalogr 26: 7-22. doi: 10.1177/155005949502600104
|
| [24] |
25. Hughes JR (1995) The phenomenon of travelling waves: A review. Clin Electroencephalogr 26:1-6. doi: 10.1177/155005949502600103
|
| [25] | 26. Amzica F, Steriade M (1995) Short- and long-range neuronal synchronization of the slow (< 1 Hz) cortical oscillation. J Neurophysiol 73: 20-38. |
| [26] | 27. Nunez PL (1995) Neocortical dynamics and human EEG rhythms. Oxford: Oxford University Press. |
| [27] |
28. Massimini M, Huber R, Ferrarelli F, et al. (2004) The sleep slow oscillation as a traveling wave. J Neurosci 24: 6862-6870. doi: 10.1523/JNEUROSCI.1318-04.2004
|
| [28] |
29. Bahramisharif A, van Gerven MA, Aarnoutse EJ, et al. (2013) Propagating neocortical gamma bursts are coordinated by traveling alpha waves. J Neurosci 33: 18849-18854. doi: 10.1523/JNEUROSCI.2455-13.2013
|
| [29] |
30. Patten TM, Rennie CJ, Robinson PA, et al. (2012) Human cortical traveling waves: Dynamical properties and correlations with responses. PLoS One 7: e38392. doi: 10.1371/journal.pone.0038392
|
| [30] |
31. Burkitt GR, Silberstein RB, Cadusch PJ, et al. (2000) Steady-state visual evoked potentials and travelling waves. Clin Neurophysiol 111: 246-258. doi: 10.1016/S1388-2457(99)00194-7
|
| [31] | 32. Pietrobon D, Moskowitz MA (2014) Chaos and commotion in the wake of cortical spreading depression and spreading depolarizations. Nat Rev Neurosci 15: 379-393. |
| [32] |
33. Kaiser D (2013) Infralow frequencies and ultradian rhythms. Semin Pediatr Neurol 20:242-245. doi: 10.1016/j.spen.2013.10.005
|
| [33] |
34. Muller L, Destexhe A (2012) Propagating waves in thalamus, cortex and the thalamocortical system: Experiments and models. J Physiol Paris 106: 222-238. doi: 10.1016/j.jphysparis.2012.06.005
|
| [34] |
35. Sato TK, Nauhaus I, Carandini M (2012) Traveling waves in visual cortex. Neuron 75: 218-229. doi: 10.1016/j.neuron.2012.06.029
|
| [35] | 36. Wu J-Y, Huang X, Zhang C (2008) Propagating waves of activity in the neocortex: What they are, what they do. Neuroscientist 14: 487-502. |
| [36] | 37. Cowey A (1964) Projection of the retina on to striate prestriate cortex in the squirrel monkey, Saimiri scireus. J Neurophysiol 27: -393. |
| [37] | 38. Grinvald A, Lieke EE, Frostig RD, et al. (1994) Cortical point-spread function and long-range lateral interactions revealed by real-time optical imaging of macaque monkey primary visual cortex. J Neurosci 14: 2545-2568. |
| [38] |
39. Horikawa J, Hosokawa Y, Kubota M, et al. (1996) Optical imaging of spatiotemporal patterns of glutamatergic excitation and GABAergic inhibition in the guinea-pig auditory cortex in vivo. J Physiol 497: 629-6 doi: 10.1113/jphysiol.1996.sp021795
|
| [39] |
40. Kubota M, Sugimoto S, Horikawa J, et al. (1997) Optical imaging of dynamic horizontal spread of excitation in rat auditory cortex slices. Neurosci Lett 237: 77-80. doi: 10.1016/S0304-3940(97)00806-9
|
| [40] |
41. Horikawa J, Hess A, Nasu M, et al. (2001) Optical imaging of neural activity in multiple auditory cortical fields of guinea pigs. Neuroreport 12: 3335-3339. doi: 10.1097/00001756-200110290-00038
|
| [41] |
42. Kubota M, Sugimoto S, Horikawa J (2008) Dynamic spatiotemporal inhibition in the guinea pig auditory cortex. Neuroreport 19: 1691-1694. doi: 10.1097/WNR.0b013e32831579ff
|
| [42] |
43. Kubota M, Miyamoto A, Hosokawa Y, et al. (2012) Spatiotemporal dynamics of neural activity related to auditory induction in the core and belt fields of guinea-pig auditory cortex. Neuroreport 23: 474-478. doi: 10.1097/WNR.0b013e328352de20
|
| [43] |
44. Grinvald A, Hildesheim R (2004) VSDI: A new era in functional imaging of cortical dynamics. Nat Rev Neurosci 5: 874-885. doi: 10.1038/nrn1536
|
| [44] |
45. Chemla S, Chavane F (2010) Voltage-sensitive dye imaging: Technique review and models. J Physiol Paris 104: 40-50. doi: 10.1016/j.jphysparis.2009.11.009
|
| [45] |
46. Mohajerani MH, McVea DA, Fingas M, et al. (2010) Mirrored bilateral slow-wave cortical activity within local circuits revealed by fast bihemispheric voltage-sensitive dye imaging in anesthetized and awake mice. J Neurosci 30: 373751. doi: 10.1523/JNEUROSCI.6437-09.2010
|
| [46] |
47. Morales-Botello ML, Aguilar J, Foffani G (2012) Imaging the spatio-temporal dynamics of supragranular activity in the rat somatosensory cortex in response to stimulation of the paws. PLoS One 7: e40174. doi: 10.1371/journal.pone.0040174
|
| [47] |
48. Ferezou I, Bolea S, Petersen CC (2006) Visualizing the cortical representation of whisker touch: Voltage-sensitive dye imaging in freely moving mice. Neuron 50: 617-629. doi: 10.1016/j.neuron.2006.03.043
|
| [48] |
49. Ferezou I, Haiss F, Gentet LJ, et al. (2007) Spatiotemporal dynamics of cortical sensorimotor integration in behaving mice. Neuron 56: 907-923. doi: 10.1016/j.neuron.2007.10.007
|
| [49] |
50. Benucci A, Frazor RA, Carandini M (2007) Standing waves and traveling waves distinguish two circuits in visual cortex. Neuron 55: 103-117. doi: 10.1016/j.neuron.2007.06.017
|
| [50] |
51. Xu W, Huang X, Takagaki K, Wu JY (2007) Compression and reflection of visually evoked cortical waves. Neuron 55: 119-129. doi: 10.1016/j.neuron.2007.06.016
|
| [51] |
52. Huang X, Xu W, Liang J, et al. (2010) Spiral wave dynamics in neocortex. Neuron 68: 978-990. doi: 10.1016/j.neuron.2010.11.007
|
| [52] |
53. Rubino D, Robbins KA, Hatsopoulos NG (2006) Propagating waves mediate information transfer in the motor cortex. Nat Neurosci 9: 1549-1557. doi: 10.1038/nn1802
|
| [53] |
54. Wanger T, Takagaki K, Lippert MT, et al. (2013) Wave propagation of cortical population activity under urethane anesthesia is state dependent. BMC Neurosci 14: 78. doi: 10.1186/1471-2202-14-78
|
| [54] |
55. Roland PE, Hanazawa A, Undeman C, et al. (2006) Cortical feedback depolarization waves: A mechanism of top-down influence on early visual areas. Proc Natl Acad Sci U S A 103:12586-12591. doi: 10.1073/pnas.0604925103
|
| [55] | 56. Muller L, Reynaud A, Chavane F, et al. (2014) The stimulus-evoked population response in visual cortex of awake monkey is a propagating wave. Nat Commun 5: 3675. |
| [56] |
57. Mohajerani MH, Chan AW, Mohsenvand M, et al. (2013) Spontaneous cortical activity alternates between motifs defined by regional axonal projections. Nat Neurosci 16: 1426-1435. doi: 10.1038/nn.3499
|
| [57] |
58. Land R, Engler G, Kral A, et al. (2012) Auditory evoked bursts in mouse visual cortex during isoflurane anesthesia. PLoS One 7: e49855. doi: 10.1371/journal.pone.0049855
|
| [58] |
59. Reimer A, Hubka P, Engel AK, et al. (2011) Fast propagating waves within the rodent auditory cortex. Cereb Cortex 21: 166-177. doi: 10.1093/cercor/bhq073
|
| [59] |
60. Han F, Caporale N, Dan Y (2008) Reverberation of recent visual experience in spontaneous cortical waves. Neuron 60: 321-327. doi: 10.1016/j.neuron.2008.08.026
|
| [60] |
61. Kral A, Tillein J, Hubka P, et al. (2009) Spatiotemporal patterns of cortical activity with bilateral cochlear implants in congenital deafness. J Neurosci 29: 811-827. doi: 10.1523/JNEUROSCI.2424-08.2009
|
| [61] |
62. Berger T, Borgdorff A, Crochet S, et al. (2007) Combined voltage and calcium epifluorescence imaging in vitro and in vivo reveals subthreshold and suprathreshold dynamics of mouse barrel cortex. J Neurophysiol 97: 3751-3762. doi: 10.1152/jn.01178.2006
|
| [62] |
63. Ermentrout GB, Kleinfeld D (2001) Traveling electrical waves in cortex: Insights from phase dynamics and speculation on a computational role. Neuron 29: 33-44. doi: 10.1016/S0896-6273(01)00178-7
|
| [63] | 64. Ermentrout B, Wang JW, Flores J, et al. (2001) Model for olfactory discrimination and learning in Limax procerebrum incorporating oscillatory dynamics and wave propagation. J Neurophysiol 85: 1444-1452. |
| [64] |
65. Kilpatrick ZP, Ermentrout B (2012) Response of traveling waves to transient inputs in neural fields. Phys Rev E Stat Nonlin Soft Matter Phys 85: 021910. doi: 10.1103/PhysRevE.85.021910
|
| [65] |
66. Heitmann S, Boonstra T, Breakspear M (2013) A dendritic mechanism for decoding traveling waves: Principles and applications to motor cortex. PLoS Comput Biol 9: e1003260. doi: 10.1371/journal.pcbi.1003260
|
| [66] |
67. Nenadic Z, Ghosh BK, Ulinski P (2003) Propagating waves in visual cortex: A large-scale model of turtle visual cortex. J Comput Neurosci 14: 161-184. doi: 10.1023/A:1021954701494
|
| [67] |
68. Wang W, Campaigne C, Ghosh BK, et al. (2005) Two cortical circuits control propagating waves in visual cortex. J Comput Neurosci 19: 263-289. doi: 10.1007/s10827-005-2288-5
|
| [68] |
69. Frohlich F, McCormick DA (2010) Endogenous electric fields may guide neocortical network activity. Neuron 67: 129-143. doi: 10.1016/j.neuron.2010.06.005
|
| [69] |
70. Silver MA, Shenhav A, D'Esposito M (2008) Cholinergic enhancement reduces spatial spread of visual responses in human early visual cortex. Neuron 60: 904-914. doi: 10.1016/j.neuron.2008.09.038
|
| [70] | 71. Meister M, Wong RO, Baylor DA, et al. (1991) Synchronous bursts of action potentials in ganglion cells of the developing mammalian retina. Science: 939-943. |
| [71] | 72. Buzsaki G (2011) Rhythms of the brain. New York: Oxford University Press. |
| [72] |
73. Blake DT, Strata F, Churchland AK, et al. (2002) Neural correlates of instrumental learning in primary auditory cortex. Proc Natl Acad Sci USA 99: 10114-10119. doi: 10.1073/pnas.092278099
|
| [73] |
74. Blake DT, Strata F, Kempter R, et al. (2005) Experience-dependent plasticity in S1 caused by noncoincident inputs. J Neurophysiol 94: 2239-2250. doi: 10.1152/jn.00172.2005
|
| [74] |
75. Blake DT, Heiser MA, Caywood M, et al. (2006) Experience-dependent adult cortical plasticity requires cognitive association between sensation and reward. Neuron 52: 371-381. doi: 10.1016/j.neuron.2006.08.009
|
| [75] |
76. Spingath EY, Kang HS, Plummer T, et al. (2011) Different neuroplasticity for task targets and distractors. PLoS One 6: e15342. doi: 10.1371/journal.pone.0015342
|
| [76] |
77. Spingath E, Kang HS, Blake DT (2013) Task-dependent modulation of SI physiological responses to targets and distractors. J Neurophysiol 109: 1036-1044. doi: 10.1152/jn.00385.2012
|
| [77] |
78. Gonzalez-Lima F, Scheich H (1986) Classical conditioning of tone-signaled bradycardia modifies 2-deoxyglucose uptake patterns in cortex, thalamus, habenula, caudate-putamen and hippocampal formation. Brain Res 363: 239-256. doi: 10.1016/0006-8993(86)91009-7
|
| [78] |
79. Hayes DJ, Duncan NW, Xu J, et al. (2014) A comparison of neural responses to appetitive and aversive stimuli in humans and other mammals. Neurosci Biobehav Rev 45: 350-368. doi: 10.1016/j.neubiorev.2014.06.018
|
| [79] |
80. Cybulska-Klosowicz A, Zakrzewska R, Kossut M (2009) Brain activation patterns during classical conditioning with appetitive or aversive UCS. Behav Brain Res 204: 102-111. doi: 10.1016/j.bbr.2009.05.024
|
| [80] |
81. McIntosh AR, Gonzalez-Lima F (1993) Network analysis of functional auditory pathways mapped with fluorodeoxyglucose: Associative effects of a tone conditioned as a Pavlovian excitor or inhibitor. Brain Res 627: 129-140. doi: 10.1016/0006-8993(93)90756-D
|
| [81] |
82. Linden JF, Liu RC, Sahani M, et al. (2003) Spectrotemporal structure of receptive fields in areas AI and AAF of mouse auditory cortex. J Neurophysiol 90: 2660-2675. doi: 10.1152/jn.00751.2002
|
| [82] |
83. David SV, Fritz JB, Shamma SA (2012) Task reward structure shapes rapid receptive field plasticity in auditory cortex. Proc Natl Acad Sci U S A 109: 2144-2149. doi: 10.1073/pnas.1117717109
|
| [83] |
84. Yin P, Fritz JB, Shamma SA (2014) Rapid spectrotemporal plasticity in primary auditory cortex during behavior. J Neurosci 34: 4396-4408. doi: 10.1523/JNEUROSCI.2799-13.2014
|
| [84] |
85. deCharms RC, Blake DT, Merzenich MM (1998) Optimizing sound features for cortical neurons. Science 280: 1439-1443. doi: 10.1126/science.280.5368.1439
|
| [85] |
86. Haider B, Duque A, Hasenstaub AR, McCormick DA (2006) Neocortical network activity in vivo is generated through a dynamic balance of excitation and inhibition. J Neurosci 26:4535-4545. doi: 10.1523/JNEUROSCI.5297-05.2006
|
| [86] |
87. Wester JC, Contreras D (2012) Columnar interactions determine horizontal propagation of recurrent network activity in neocortex. J Neurosci 32: 5454-5471. doi: 10.1523/JNEUROSCI.5006-11.2012
|
| [87] | 88. O'Reilly RC (2001) Generalization in interactive networks: The benefits of inhibitory competition and Hebbian learning. Neural Comput 13: 11991-11241. |
| [88] |
89. Swindale NV (2004) How different feature spaces may be represented in cortical maps. Network: Comput Neural Syst 15: 217-242. doi: 10.1088/0954-898X/15/4/001
|
| [89] | 90. Roux L, Buzsaki G (2014) Tasks for inhibitory interneurons in intact brain circuits. Neuropharmacology. |
| [90] | 91. Brunel N, Wang XJ (2003) What determines the frequency of fast network oscillations with irregular neural discharges? I. Synaptic dynamics and excitation-inhibition balance. J Neurophysiol 415-430. |
| [91] |
92. Orduña I, Mercado E, III, Gluck MA, et al. (2005) Cortical responses in rats predict perceptual sensitivities to complex sounds. Behav Neurosci 119: 256-264. doi: 10.1037/0735-7044.119.1.256
|
| [92] |
93. Engineer CT, Perez CA, Chen YH, et al. (2008) Cortical activity patterns predict speech discrimination ability. Nat Neurosci 11: 603-608. doi: 10.1038/nn.2109
|
| [93] |
94. Engineer CT, Perez CA, Carraway RS, et al. (2013) Similarity of cortical activity patterns predicts generalization behavior. PLoS One 8: e78607. doi: 10.1371/journal.pone.0078607
|
| [94] |
95. McLin DE, III, Miasnikov AA, Weinberger NM (2003) CS-specific gamma, theta, and alpha EEG activity detected in stimulus generalization following induction of behavioral memory by stimulation of the nucleus basalis. Neurobiol Learn Mem 79: 152-176. doi: 10.1016/S1074-7427(02)00009-6
|
| [95] | 96. Thompson RF (1965) The neural basis of stimulus generalization. In: Mostofsky DI, editor. Stimulus generalization. Stanford: Stanford University Press, 154-178. |
| [96] |
97. Mercado E, III, Orduña I, Nowak JM (2005) Auditory categorization of complex sounds by rats (Rattus norvegicus). J Comp Psychol 119: 90-98. doi: 10.1037/0735-7036.119.1.90
|
| [97] |
98. Weisman RG, Njegovan MG, Williams MT, et al. (2004). A behavior analysis of absolute pitch: Sex, experience, and species. Behav Processes 66: 289-307. doi: 10.1016/j.beproc.2004.03.010
|
| [98] |
99. Wisniewski MG, Church BA, Mercado E, III (2009) Learning-related shifts in generalization gradients for complex sounds. Learn Behav 37: 325-335. doi: 10.3758/LB.37.4.325
|
| [99] |
100. Wisniewski MG, Church BA, Mercado E, III (2014) Individual differences during acquisition predict shifts in generalization. Behav Processes 104: 26-34. doi: 10.1016/j.beproc.2014.01.007
|
| [100] |
101. Church BA, Mercado E, III, Wisniewski MG, et al (2013) Temporal dynamics in auditory perceptual learning: Impact of sequencing and incidental learning. J Exp Psychol Learn Mem Cogn 39: 270-276. doi: 10.1037/a0028647
|
| [101] |
102. Liu EH, Mercado E, III, Church BA, et al. (2008) The easy-to-hard effect in human (Homo sapiens) and rat (Rattus norvegicus) auditory identification. J Comp Psychol 122: 132-145. doi: 10.1037/0735-7036.122.2.132
|
| [102] |
103. Schreiner CE, Polley DB (2014) Auditory map plasticity: Diversity in causes and consequences. Curr Opin Neurobiol 24: 143-156. doi: 10.1016/j.conb.2013.11.009
|
| [103] |
104. Mercado E, III (2008) Neural and cognitive plasticity: From maps to minds. Psychol Bull 134:109-137. doi: 10.1037/0033-2909.134.1.109
|
| [104] | 105. Mercado E, III (2011) Mapping individual variations in learning capacity. Int J Comp Psychol24: 4-35. |
| [105] |
106. Ghirlanda S, Enquist M (2003) A century of generalization. Anim Behav 66: 15-36. doi: 10.1006/anbe.2003.2174
|
| [106] |
107. McLaren IP, Mackintosh NJ (2002) Associative learning and elemental representation: II. Generalization and discrimination. Anim Learn Behav 30: 177-200. doi: 10.3758/BF03192828
|
| [107] |
108. Lazareva OF, Young ME, Wasserman EA (2014) A three-component model of relational responding in the transposition paradigm. J Exp Psychol Anim Learn Cogn 40: 63-80. doi: 10.1037/xan0000004
|
| [108] |
109. Wisniewski MG, Radell ML, Guillette LM, et al. (2012) Predicting shifts in generalization gradients with perceptrons. Learn Behav 40: 128-144. doi: 10.3758/s13420-011-0050-6
|
| [109] |
110. Litaudon P, Mouly AM, Sullivan R, et al. (1997) Learning-induced changes in rat piriform cortex activity mapped using multisite recording with voltage sensitive dye. Eur J Neurosci 9:1593-1602. doi: 10.1111/j.1460-9568.1997.tb01517.x
|
| [110] | 111. Basar E (2004) Memory and brain dynamics: Oscillations integrating attention, perception, learning and memory. Boca Raton: CRC Press. |
| [111] |
112. Orduña I, Liu EH, Church BA, et al. (2012) Evoked-potential changes following discrimination learning involving complex sounds. Clin Neurophysiol 123: 711-719. doi: 10.1016/j.clinph.2011.08.019
|
| [112] |
113. Freyer F, Becker R, Dinse HR, et al. (2013) State-dependent perceptual learning. J Neurosci 33:2900-2907. doi: 10.1523/JNEUROSCI.4039-12.2013
|
| [113] | 114. Kanai R, Rees G (2011) The structural basis of inter-individual differences in human behaviour and cognition. Nat Rev Neurosci 12: 231-242. |
| [114] |
115. Buzsaki G (2005) Theta rhythm of navigation: Link between path integration and landmark navigation, episodic and semantic memory. Hippocampus 15: 827-840. doi: 10.1002/hipo.20113
|
| [115] | 116. Hasselmo ME (2005) What is the function of hippocampal theta rhythm?—Linking behavioral data to phasic properties of field potential and unit recording data. Hippocampus 15: 936-949. |
| [116] |
117. Patel J, Fujisawa S, Berenyi A, et al. (2012) Traveling theta waves along the entire septotemporal axis of the hippocampus. Neuron 75: 410-417. doi: 10.1016/j.neuron.2012.07.015
|
| [117] | 118. Lubenov EV, Siapas AG (2009) Hippocampal theta oscillations are travelling waves. Nature459: 534-539. |
| [118] |
119. Tsanov M, Wright N, Vann SD, et al. (2011) Hippocampal inputs mediate theta-related plasticity in anterior thalamus. Neuroscience 187: 52-62. doi: 10.1016/j.neuroscience.2011.03.055
|
| [119] |
120. Hoffmann LC, Berry SD (2009) Cerebellar theta oscillations are synchronized during hippocampal theta-contingent trace conditioning. Proc Natl Acad Sci U S A 106: 21371-21376. doi: 10.1073/pnas.0908403106
|
| [120] |
121. Darling RD, Takatsuki K, Griffin AL, et al. (2011) Eyeblink conditioning contingent on hippocampal theta enhances hippocampal and medial prefrontal responses. J Neurophysiol 105:2213-2224. doi: 10.1152/jn.00801.2010
|
| [121] | 122. Hyman JM, Hasselmo ME, Seamans JK (2011) What is the functional relevance of prefrontal cortex entrainment to hippocampal theta rhythms? Front Neurosci 5: 24. |
| [122] |
123. Seager MA, Johnson LD, Chabot ES, et al. (2002) Oscillatory brain states and learning: Impact of hippocampal theta-contingent training. Proc Natl Acad Sci U S A 99: 1616-1620. doi: 10.1073/pnas.032662099
|
| [123] |
124. Griffin AL, Asaka Y, Darling RD, et al. (2004) Theta-contingent trial presentation accelerates learning rate and enhances hippocampal plasticity during trace eyeblink conditioning. Behav Neurosci 118: 403-411. doi: 10.1037/0735-7044.118.2.403
|
| [124] |
125. Berry SD, Hoffmann LC (2011) Hippocampal theta-dependent eyeblink classical conditioning: Coordination of a distributed learning system. Neurobiol Learn Mem 95: 185-189. doi: 10.1016/j.nlm.2010.11.014
|
| [125] |
126. Burke JF, Sharan AD, Sperling MR, et al. (2014) Theta and high-frequency activity mark spontaneous recall of episodic memories. J Neurosci 34: 11355-11365. doi: 10.1523/JNEUROSCI.2654-13.2014
|
| [126] | 127. Squire LR, Kandel ER (1999) Memory: From minds to molecules. New York: Scientific American Library. |
| [127] |
128. Cabeza R, Moscovitch M (2013) Memory systems, processing modes, and components: Functional neuroimaging evidence. Perspect Psychol Sci 8: 49-55. doi: 10.1177/1745691612469033
|
| [128] | 129. Craik FIM (1985) Paradigms in human memory research. In: Nilsson L-G, Archer T, editors. Perspectives on learning and memory. Hillsdale, NY: Lawrence Erlbaum, 197-221. |
| [129] | 130. Capaldi EJ, Neath I (1995) Remembering and forgetting as context discrimination. Learn Mem2: 107-132. |
| [130] |
132. Neath I, Crowder RG (1990) Schedules of presentation and temporal distinctiveness in human memory. J Exp Psychol Learn Mem Cogn 16: 316-327. doi: 10.1037/0278-7393.16.2.316
|
| [131] |
133. Neath I (1993) Distinctiveness and serial position effects in recognition. Mem Cognit 21:689-698. doi: 10.3758/BF03197199
|
| [132] |
134. Nairne JS (1991) Positional uncertainty in long-term memory. Mem Cognit 19: 332-340. doi: 10.3758/BF03197136
|
| [133] |
135. Wright AA (1999) Auditory list memory and interference processes in monkeys. J Exp Psychol Anim Behav Process 25: 284-296. doi: 10.1037/0097-7403.25.3.284
|
| [134] |
136. Wright AA, Roediger HL, III (2003) Interference processes in monkey auditory list memory. Psychon Bull Rev 10: 696-702. doi: 10.3758/BF03196534
|
| [135] | 137. Kosslyn SM, Ganis G, Thompson WL (2001) Neural foundations of imagery. Nat Rev Neurosci2: 635-642. |
| [136] |
138. Euston DR, Tatsuno M, McNaughton BL (2007) Fast-forward playback of recent memory sequences in prefrontal cortex during sleep. Science 318: 1147-1150. doi: 10.1126/science.1148979
|
| [137] |
139. McVea DA, Mohajerani MH, Murphy TH (2012) Voltage-sensitive dye imaging reveals dynamic spatiotemporal properties of cortical activity after spontaneous muscle twitches in the newborn rat. J Neurosci 32: 10982-10994. doi: 10.1523/JNEUROSCI.1322-12.2012
|
| [138] | 140. Draaisma D (2000) Metaphors of memory: A history of ideas about the mind. Cambridge: Cambridge University Press. |
| [139] | 141. Rudy JW (2014) The neurobiology of learning and memory. Sunderland, MA: Sinauer Associates. |
| [140] |
142. Lee H, Fell J, Axmacher N (2013) Electrical engram: How deep brain stimulation affects memory. Trends Cogn Sci 17: 574-584. doi: 10.1016/j.tics.2013.09.002
|
| [141] |
143. Bartolomei F, Barbeau E, Gavaret M, et al. (2004) Cortical stimulation study of the role of rhinal cortex in deja vu and reminiscence of memories. Neurology 63: 858-864. doi: 10.1212/01.WNL.0000137037.56916.3F
|
| [142] |
144. Bartolomei F, Barbeau EJ, Nguyen T, et al. (2012) Rhinal-hippocampal interactions during deja vu. Clin Neurophysiol 123: 489-495. doi: 10.1016/j.clinph.2011.08.012
|
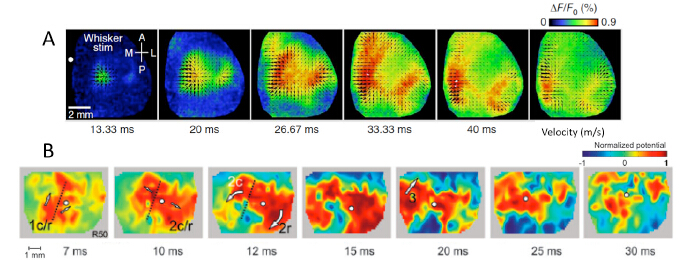
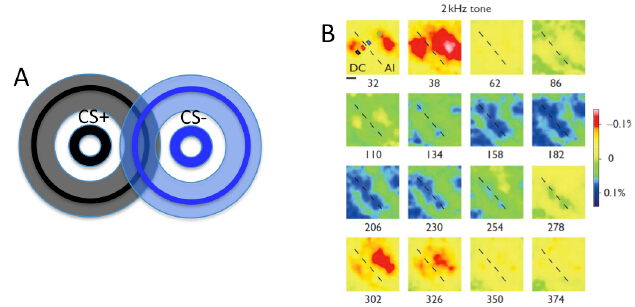
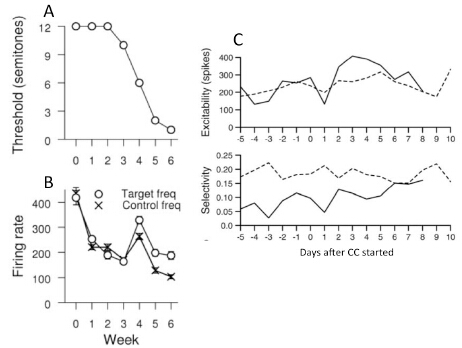
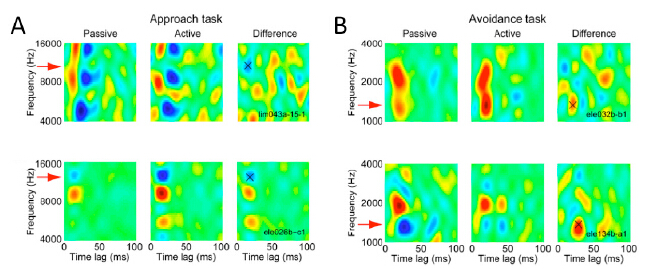
Figures(4)

Eduardo Mercado III. Relating Cortical Wave Dynamics to Learning and Remembering[J]. AIMS Neuroscience, 2014, 1(3): 185-209. doi: 10.3934/Neuroscience.2014.3.185







 DownLoad:
DownLoad: