Citation: Leena Latonen. Protein aggregation in neurodegenerative disease: the nucleolar connection[J]. AIMS Molecular Science, 2015, 2(3): 324-331. doi: 10.3934/molsci.2015.3.324

| [1] | Labbadia J, Morimoto RI (2015) The Biology of Proteostasis in Aging and Disease. Annu Rev Biochem 84: 435-464. |
| [2] |
Gallagher PS, Oeser ML, Abraham AC, et al. (2014) Cellular maintenance of nuclear protein homeostasis. Cell Mol Life Sci 71: 1865-1879. doi: 10.1007/s00018-013-1530-y

|
| [3] |
Hart FU, Bracher A, Hayer-Hartl M (2011) Molecular chaperones in protein folding and proteostasis. Nature 475: 324-332. doi: 10.1038/nature10317
|
| [4] | Lamark T, Johansen T (2012) Aggrephagy: selective disposal of protein aggregates by macroautophagy. Int J Cell Biol 2012: 736905. |
| [5] | Shibata Y, Morimoto RI (2014) How the nucleus copes with proteotoxic stress. CurrBiol 24: R463-474. |
| [6] | Gray DA, Woulfe J (2013) Structural disorder and the loss of RNA homeostasis in aging and neurodegenerative disease. Front Genet 4: 149. |
| [7] |
Arslan MA, Chikina M, Csermely P, et al. (2012) Misfolded proteins inhibit proliferation and promote stress-induced death in SV40-transformed mammalian cells. FASEB J 26: 766-777. doi: 10.1096/fj.11-186197
|
| [8] |
Bence NF, Sampat RM, Kopito RR (2001) Impairment of the ubiquitin-proteasome system by protein aggregation. Science 292: 1552-1555. doi: 10.1126/science.292.5521.1552
|
| [9] |
Holmberg CI, Staniszewski KE, Mensah KN, et al. (2004) Inefficient degradation of truncated polyglutamine proteins by the proteasome. EMBO J 23: 4307-4318. doi: 10.1038/sj.emboj.7600426
|
| [10] |
Bennett EJ, Bence NF, Jayakumar R, et al. (2005) Global impairment of the ubiquitin-proteasome system by nuclear or cytoplasmic protein aggregates precedes inclusion body formation. Mol Cell 17: 351-365. doi: 10.1016/j.molcel.2004.12.021
|
| [11] |
Hipp MS, Patel CN, Bersuker K, et al. (2012) Indirect inhibition of 26S proteasome activity in a cellular model of Huntington’s disease. J Cell Biol 196: 573-857. doi: 10.1083/jcb.201110093
|
| [12] |
Hipp MS, Park SH, Hartl FU (2014) Proteostasis impairment in protein-misfolding and -aggregation diseases. Trends Cell Biol 24: 506-514. doi: 10.1016/j.tcb.2014.05.003
|
| [13] | Takalo M, Salminen A, Soininen H, et al. (2013) Protein aggregation and degradation mechanisms in neurodegenerative diseases. Am J Neurodegener Dis 2: 1-14. |
| [14] |
Latonen L (2011) Nucleolar aggresomes as counterparts of cytoplasmic aggresomes in proteotoxic stress. Bioessays 33: 386-395. doi: 10.1002/bies.201100008
|
| [15] | Woulfe J (2008) Nuclear bodies in neurodegenerative disease. Biochim BiophysActa 1783: 2195-2206. |
| [16] |
Ramaswami M, Taylor JP, Parker R (2013) RNA-Protein Granules in Degenerative Disorders. Cell 154: 727-736. doi: 10.1016/j.cell.2013.07.038
|
| [17] |
Miller SB, Ho CT, Winkler J, et al. (2015) Compartment-specific aggregases direct distinct nuclear and cytoplasmic aggregate deposition. EMBO J 34: 778-797. doi: 10.15252/embj.201489524
|
| [18] |
Dundr M (2012) Nuclear bodies: multifunctional companions of the genome. Curr Opin Cell Biol 24: 415-422. doi: 10.1016/j.ceb.2012.03.010
|
| [19] |
Kaganovich D, Kopito R, Frydman J (2008) Misfolded proteins partition between two distinct quality control compartments. Nature 454: 1088-1095. doi: 10.1038/nature07195
|
| [20] |
Park SH, Kukushkin Y, Gupta R, et al. (2013) PolyQ proteins interfere with nuclear degradation of cytosolic proteins by sequestering the Sis1p chaperone. Cell 154: 134-145. doi: 10.1016/j.cell.2013.06.003
|
| [21] | Lu M, Williamson N, Boschetti C, et al. (2015) Expression-level dependent perturbation of cell proteostasis and nuclear morphology by aggregation-prone polyglutamine proteins. Biotechnol Bioeng. doi: 10.1002/bit.25606. |
| [22] |
Caron NS, Hung CL, Atwal RS, et al. (2014) Live cell imaging and biophotonic methods reveal two types of mutant huntingin inclusions. Hum Mol Genet 23: 2324-2338. doi: 10.1093/hmg/ddt625
|
| [23] |
Chapple JP, Bros-Facer V, Butler R, et al. (2008) Focal distortion of the nuclear envelope by huntingtin aggregates revealed by lamin immunostaining. Neurosci Lett 447: 172-174. doi: 10.1016/j.neulet.2008.09.075
|
| [24] |
Mapelli L, Canale C, Pesci D, et al. (2012) Toxic effects of expanded ataxin-1 involve mechanical instability of the nuclear membrane. Biochim Biophys Acta 1822: 906-917. doi: 10.1016/j.bbadis.2012.01.016
|
| [25] | Lindström MS, Latonen L (2013) The nucleolus as a stress respose organelle. In: Proteins of the Nucleolus. Regulation, Translocation and Biomedical Functions. Springer, 251-273. |
| [26] |
Andersen JS, Lam YW, Leung AK, et al. (2005) Nucleolar proteome dynamics. Nature 433: 77-83. doi: 10.1038/nature03207
|
| [27] |
Latonen L, Moore HM, Bai B, et al. (2011) Proteasome inhibitors induce nucleolar aggregation of proteasome target proteins and polyadenylated RNA by altering ubiquitin availability. Oncogene 30: 790-805. doi: 10.1038/onc.2010.469
|
| [28] |
Krüger T, Scheer U (2010) p53 localizes to intranucleolar regions distinct from the ribosome production compartments. J Cell Sci 123: 1203-1208. doi: 10.1242/jcs.062398
|
| [29] |
Vilotti S, Biagioli M, Foti R, et al. (2012) The PML nuclear bodies-associated protein TTRAP regulates ribosome biogenesis in nucleolar cavities upon proteasome inhibition. Cell Death Differ 19: 488-500. doi: 10.1038/cdd.2011.118
|
| [30] |
Ehm P, Nalaskowski MM, Wundenberg T, et al. (2015) The tumor suppressor SHIP1 colocalizes in nucleolar cavities with p53 and components of PML nuclear bodies. Nucleus 6: 154-164. doi: 10.1080/19491034.2015.1022701
|
| [31] |
Nollen EA, Salomons FA, Brunsting JF, et al. (2001) Dynamic changes in the localization of thermally unfolded nuclear proteins associated with chaperone-dependent protection. Proc Natl AcadSci USA 98: 12038-12043. doi: 10.1073/pnas.201112398
|
| [32] |
Forsberg K, Andersen PM, Marklund SL, et al. (2011) Glial nuclear aggregates of superoxide dismutase-1 are regularly present in patients with amyotrophic lateral sclerosis. Acta Neuropathol 121: 623-634. doi: 10.1007/s00401-011-0805-3
|
| [33] | Li M, Nakagomi Y, Kobayashi Y, et al. (1998) Nonneural nuclear inclusions of androgen receptor protein in spinal and bulbar muscular atrophy. Am J Pathol 153: 695-701. |
| [34] |
Hetman M, Pietrzak M (2012) Emerging roles of the neuronal nucleolus. Trends Neurosci 35: 305-314. doi: 10.1016/j.tins.2012.01.002
|
| [35] |
Parlato R, Kreiner G (2013) Nucleolar activity in neurodegenerative diseases: a missing piece of the puzzle? J Mol Med (Berl) 91: 541-547. doi: 10.1007/s00109-012-0981-1
|
| [36] |
Stavreva DA, Kawasaki M, Dundr M, et al. (2006) Potential roles for ubiquitin and the proteasome during ribosome biogenesis. Mol Cell Biol 26: 5131-5145 doi: 10.1128/MCB.02227-05
|
| [37] |
Nyström T, Liu B (2014) The mystery of aging and rejuvenation—a budding topic. Curr Opin Microbiol 18: 61-67. doi: 10.1016/j.mib.2014.02.003
|
| [38] |
Song J, Yang Q, Yang J, et al. (2014) Essential genetic interactors of SIR2 required for spatial sequestration and asymmetrical inheritance of protein aggregates. PLoS Genet 10: e1004539. doi: 10.1371/journal.pgen.1004539
|
| [39] |
Liu B, Larsson L, Caballero A, et al. (2010) The polarisome is required for segregation and retrograde transport of protein aggregates. Cell 140: 257-267. doi: 10.1016/j.cell.2009.12.031
|
| [40] |
Zhou C, Slaughter BD, Unruh JR, et al. (2014) Organelle-based aggregation and retention of damaged proteins in asymmetrically dividing cells. Cell 159: 530-542. doi: 10.1016/j.cell.2014.09.026
|
| [41] | Katajisto P, Döhla J, Chaffer CL, et al. (2015) Stem cells. Asymmetric apportioning of aged mitochondria between daughter cells is required for stemness. Science 348: 340-343. |
| [42] |
Mogk A, Bukau B (2014) Mitochondria tether protein trash to rejuvenate cellular environments. Cell 159: 471-472. doi: 10.1016/j.cell.2014.10.007
|
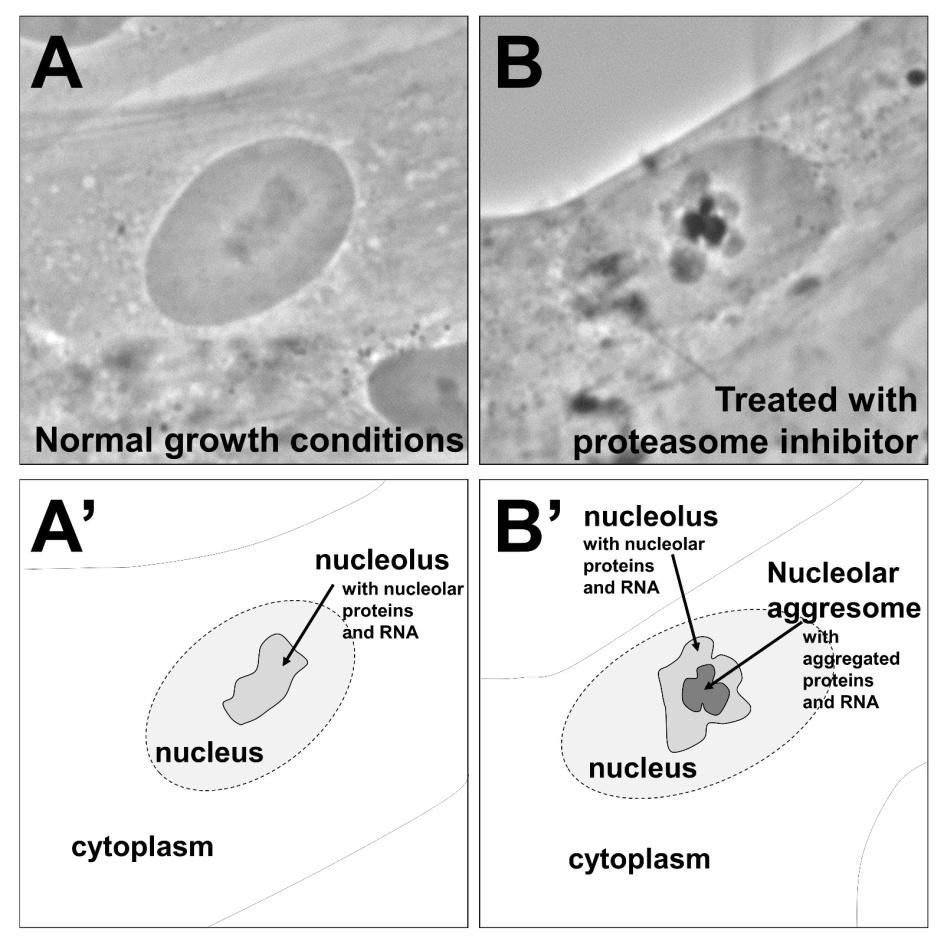
Figures(1)

Leena Latonen. Protein aggregation in neurodegenerative disease: the nucleolar connection[J]. AIMS Molecular Science, 2015, 2(3): 324-331. doi: 10.3934/molsci.2015.3.324







 DownLoad:
DownLoad: