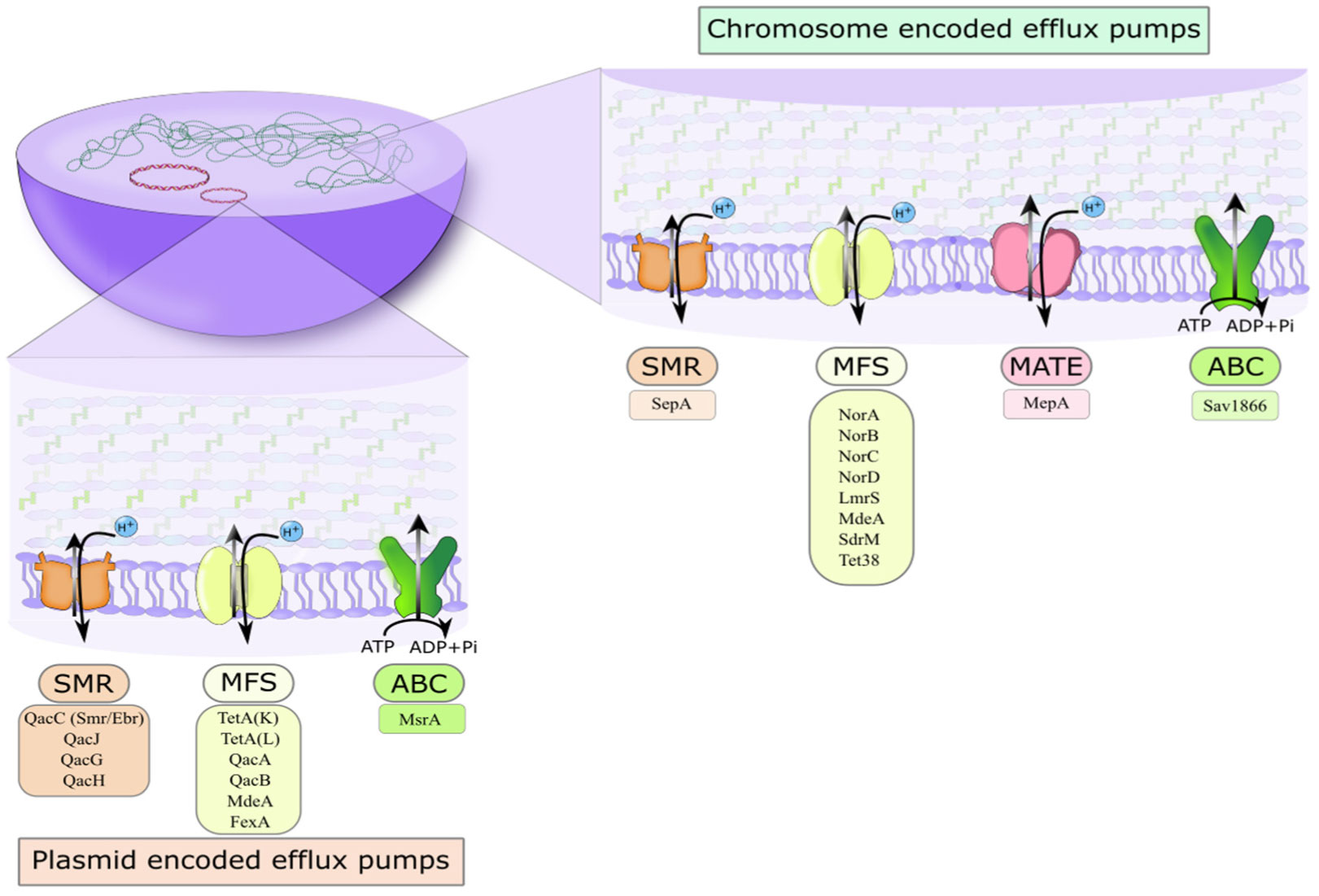
The microorganism Staphylococcus aureus is a notorious causative agent of bacterial infection. The widespread presence of this pathogen has caused significant morbidity and mortality rates in clinical healthcare settings and communities. Due to its increasingly frequent recalcitrant nature towards clinically available antimicrobial agents, the bacterium poses a considerable public health crisis. A significant bacterial mechanism of antimicrobial agent resistance includes multidrug efflux pump systems. These antimicrobial efflux determinants translate into several large superfamilies of transporters that share related amino acid sequences, similarities in three-dimensional structures, modes of energization, and solute transport catalysis across the membrane. Because of their ubiquitous nature and functional role in virulence, these multidrug transporters make good targets for inhibition. This review briefly summarizes recent key findings regarding multidrug efflux activity and modulation in the MATE, SMR, and MFS transporters.
Citation: Manjusha Lekshmi, Jerusha Stephen, Manisha Ojha, Sanath Kumar, Manuel Varela. Staphylococcus aureus antimicrobial efflux pumps and their inhibitors: recent developments[J]. AIMS Medical Science, 2022, 9(3): 367-393. doi: 10.3934/medsci.2022018
The microorganism Staphylococcus aureus is a notorious causative agent of bacterial infection. The widespread presence of this pathogen has caused significant morbidity and mortality rates in clinical healthcare settings and communities. Due to its increasingly frequent recalcitrant nature towards clinically available antimicrobial agents, the bacterium poses a considerable public health crisis. A significant bacterial mechanism of antimicrobial agent resistance includes multidrug efflux pump systems. These antimicrobial efflux determinants translate into several large superfamilies of transporters that share related amino acid sequences, similarities in three-dimensional structures, modes of energization, and solute transport catalysis across the membrane. Because of their ubiquitous nature and functional role in virulence, these multidrug transporters make good targets for inhibition. This review briefly summarizes recent key findings regarding multidrug efflux activity and modulation in the MATE, SMR, and MFS transporters.

| [1] |
Tong SY, Davis JS, Eichenberger E, et al. (2015) Staphylococcus aureus infections: epidemiology, pathophysiology, clinical manifestations, and management. Clin Microbiol Rev 28: 603-661. https://doi.org/10.1128/CMR.00134-14 
|
| [2] |
Newsom SW (2008) Ogston's coccus. J Hosp Infect 70: 369-372. https://doi.org/10.1016/j.jhin.2008.10.001
|
| [3] |
Andersen JL, He GX, Kakarla P, et al. (2015) Multidrug efflux pumps from Enterobacteriaceae, Vibrio cholerae and Staphylococcus aureus bacterial food pathogens. Int J Environ Res Public Health 12: 1487-1547. https://doi.org/10.3390/ijerph120201487
|
| [4] |
Turner NA, Sharma-Kuinkel BK, Maskarinec SA, et al. (2019) Methicillin-resistant Staphylococcus aureus: an overview of basic and clinical research. Nat Rev Microbiol 17: 203-218. https://doi.org/10.1038/s41579-018-0147-4
|
| [5] |
Murray CJL, Ikuta KS, Sharara F, et al. (2022) Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 399: 629-655. https://doi.org/10.1016/S0140-6736(21)02724-0
|
| [6] |
Varela MF, Wilson TH (1996) Molecular biology of the lactose carrier of Escherichia coli. Biochim Biophys Acta 1276: 21-34. https://doi.org/10.1016/0005-2728(96)00030-8
|
| [7] |
Kumar S, Lekshmi M, Parvathi A, et al. (2017) Antibiotic resistance in seafood borne pathogens. Foodborne Pathogens and Antibiotic Resistance . Wiley-Blackwell 397-415. https://doi.org/10.1002/9781119139188.ch17
|
| [8] |
Blair J, Webber MA, Baylay AJ, et al. (2015) Molecular mechanisms of antibiotic resistance. Nat Rev Microbiol 13: 42-51. https://doi.org/10.1038/nrmicro3380
|
| [9] |
Aslam B, Wang W, Arshad MI, et al. (2018) Antibiotic resistance: a rundown of a global crisis. Infect Drug Resist 11: 1645-1658. https://doi.org/10.2147/IDR.S173867
|
| [10] |
Lekshmi M, Ammini P, Adjei J, et al. (2018) Modulation of antimicrobial efflux pumps of the major facilitator superfamily in Staphylococcus aureus. AIMS Microbiol 4: 1-18. https://doi.org/10.3934/microbiol.2018.1.1
|
| [11] |
Varela MF, Stephen J, Lekshmi M, et al. (2021) Bacterial resistance to antimicrobial agents. Antibiotics (Basel) 10: 593. https://doi.org/10.3390/antibiotics10050593
|
| [12] |
Vestergaard M, Frees D, Ingmer H (2019) Antibiotic resistance and the MRSA problem. Microbiol Spectr 7. https://doi.org/10.1128/microbiolspec.GPP3-0057-2018
|
| [13] |
Pantosti A, Sanchini A, Monaco M (2007) Mechanisms of antibiotic resistance in Staphylococcus aureus. Future Microbiol 2: 323-334. https://doi.org/10.2217/17460913.2.3.323
|
| [14] |
Sabath LD, Finland M (1962) Inactivation of methicillin, oxacillin and ancillin by Staphylococcus aureus. Proc Soc Exp Biol Med 111: 547-550. https://doi.org/10.3181/00379727-111-27850
|
| [15] |
Frère JM, Sauvage E, Kerff F (2016) From “An Enzyme Able to Destroy Penicillin” to Carbapenemases: 70 years of β-lactamase misbehaviour. Curr Drug Targets 17: 974-982. https://doi.org/10.2174/1389450116666151001112859
|
| [16] |
Rodriguez-Bano J, Pascual A (2008) Clinical significance of extended-spectrum β-lactamases. Expert Rev Anti Infect Ther 6: 671-683. https://doi.org/10.1586/14787210.6.5.671
|
| [17] |
Sawa T, Kooguchi K, Moriyama K (2020) Molecular diversity of extended-spectrum β-lactamases and carbapenemases, and antimicrobial resistance. J Intensive Care 8: 13. https://doi.org/10.1186/s40560-020-0429-6
|
| [18] |
Lee S, Mir RA, Park SH, et al. (2020) Prevalence of extended-spectrum β-lactamases in the local farm environment and livestock: challenges to mitigate antimicrobial resistance. Crit Rev Microbiol 46: 1-14. https://doi.org/10.1080/1040841X.2020.1715339
|
| [19] |
Bayer AS, Schneider T, Sahl HG (2013) Mechanisms of daptomycin resistance in Staphylococcus aureus: role of the cell membrane and cell wall. Ann N Y Acad Sci 1277: 139-158. https://doi.org/10.1111/j.1749-6632.2012.06819.x
|
| [20] |
Guo Y, Pfahler NM, Volpel SL, et al. (2021) Cell wall glycosylation in Staphylococcus aureus: targeting the tar glycosyltransferases. Curr Opin Struct Biol 68: 166-174. https://doi.org/10.1016/j.sbi.2021.01.003
|
| [21] |
Shahid AH, Nazir K, El Zowalaty ME, et al. (2021) Molecular detection of vancomycin and methicillin resistance in Staphylococcus aureus isolated from food processing environments. One Health 13: 100276. https://doi.org/10.1016/j.onehlt.2021.100276
|
| [22] |
Lambert PA (2005) Bacterial resistance to antibiotics: modified target sites. Adv Drug Deliv Rev 57: 1471-1485. https://doi.org/10.1016/j.addr.2005.04.003
|
| [23] |
Badshah SL, Ullah A (2018) New developments in non-quinolone-based antibiotics for the inhibition of bacterial gyrase and topoisomerase IV. Eur J Med Chem 152: 393-400. https://doi.org/10.1016/j.ejmech.2018.04.059
|
| [24] |
Tremblay S, Lau TT, Ensom MH (2013) Addition of rifampin to vancomycin for methicillin-resistant Staphylococcus aureus infections: what is the evidence?. Ann Pharmacother 47: 1045-1054. https://doi.org/10.1345/aph.1R726
|
| [25] |
Shanson DC (1981) Antibiotic-resistant Staphylococcus aureus. J Hosp Infect 2: 11-36. https://doi.org/10.1016/0195-6701(81)90003-7
|
| [26] |
Nguyen F, Starosta AL, Arenz S, et al. (2014) Tetracycline antibiotics and resistance mechanisms. Biol Chem 395: 559-575. https://doi.org/10.1515/hsz-2013-0292
|
| [27] |
Liu WT, Chen EZ, Yang L, et al. (2021) Emerging resistance mechanisms for 4 types of common anti-MRSA antibiotics in Staphylococcus aureus: A comprehensive review. Microb Pathog 156: 104915. https://doi.org/10.1016/j.micpath.2021.104915
|
| [28] |
Dantley KA, Dannelly HK, Burdett V (1998) Binding interaction between Tet(M) and the ribosome: requirements for binding. J Bacteriol 180: 4089-4092. https://doi.org/10.1128/JB.180.16.4089-4092.1998
|
| [29] |
Burdett V (1996) Tet(M)-promoted release of tetracycline from ribosomes is GTP dependent. J Bacteriol 178: 3246-3251. https://doi.org/10.1128/jb.178.11.3246-3251.1996
|
| [30] |
Burdett V (1991) Purification and characterization of Tet(M), a protein that renders ribosomes resistant to tetracycline. J Biol Chem 266: 2872-2877. https://doi.org/10.1016/S0021-9258(18)49928-0
|
| [31] | Lewis K (2008) Multidrug tolerance of biofilms and persister cells. Curr Top Microbiol Immunol 322: 107-131. https://doi.org/10.1007/978-3-540-75418-3_6 |
| [32] |
Elkhatib WF, Khairalla AS, Ashour HM (2014) Evaluation of different microtiter plate-based methods for the quantitative assessment of Staphylococcus aureus biofilms. Future Microbiol 9: 725-735. https://doi.org/10.2217/fmb.14.33
|
| [33] |
Smith KP, Kumar S, Varela MF (2009) Identification, cloning, and functional characterization of EmrD-3, a putative multidrug efflux pump of the major facilitator superfamily from Vibrio cholerae O395. Arch Microbiol 191: 903-911. https://doi.org/10.1007/s00203-009-0521-8
|
| [34] | Kumar S, Varela MF (2013) Molecular mechanisms of bacterial resistance to antimicrobial agents. Microbial pathogens and strategies for combating them: science, technology and education . Formatex Research Center 522-534. |
| [35] |
Velamakanni S, Yao Y, Gutmann DA, et al. (2008) Multidrug transport by the ABC transporter Sav1866 from Staphylococcus aureus. Biochemistry 47: 9300-9308. https://doi.org/10.1021/bi8006737
|
| [36] |
Orelle C, Mathieu K, Jault JM (2019) Multidrug ABC transporters in bacteria. Res Microbiol 170: 381-391. https://doi.org/10.1016/j.resmic.2019.06.001
|
| [37] |
Forrest LR, Kramer R, Ziegler C (2011) The structural basis of secondary active transport mechanisms. Biochim Biophys Acta 1807: 167-188. https://doi.org/10.1016/j.bbabio.2010.10.014
|
| [38] |
Kuroda T, Tsuchiya T (2009) Multidrug efflux transporters in the MATE family. Biochim Biophys Acta 1794: 763-768. https://doi.org/10.1016/j.bbapap.2008.11.012
|
| [39] |
Moriyama Y, Hiasa M, Matsumoto T, et al. (2008) Multidrug and toxic compound extrusion (MATE)-type proteins as anchor transporters for the excretion of metabolic waste products and xenobiotics. Xenobiotica 38: 1107-1118. https://doi.org/10.1080/00498250701883753
|
| [40] |
Kusakizako T, Claxton DP, Tanaka Y, et al. (2019) Structural basis of H+-dependent conformational change in a bacterial MATE transporter. Structure 27: 293-301.e3. https://doi.org/10.1016/j.str.2018.10.004
|
| [41] |
Brown MH, Paulsen IT, Skurray RA (1999) The multidrug efflux protein NorM is a prototype of a new family of transporters. Mol Microbiol 31: 394-395. https://doi.org/10.1046/j.1365-2958.1999.01162.x
|
| [42] |
He X, Szewczyk P, Karyakin A, et al. (2010) Structure of a cation-bound multidrug and toxic compound extrusion transporter. Nature 467: 991-994. https://doi.org/10.1038/nature09408
|
| [43] |
Lu M, Symersky J, Radchenko M, et al. (2013) Structures of a Na+-coupled, substrate-bound MATE multidrug transporter. Proc Natl Acad Sci U S A 110: 2099-2104. https://doi.org/10.1073/pnas.1219901110
|
| [44] |
Lu M, Radchenko M, Symersky J, et al. (2013) Structural insights into H+-coupled multidrug extrusion by a MATE transporter. Nat Struct Mol Biol 20: 1310-1317. https://doi.org/10.1038/nsmb.2687
|
| [45] |
Kusakizako T, Miyauchi H, Ishitani R, et al. (2020) Structural biology of the multidrug and toxic compound extrusion superfamily transporters. Biochim Biophys Acta Biomembr 1862: 183154. https://doi.org/10.1016/j.bbamem.2019.183154
|
| [46] |
McAleese F, Petersen P, Ruzin A, et al. (2005) A novel MATE family efflux pump contributes to the reduced susceptibility of laboratory-derived Staphylococcus aureus mutants to tigecycline. Antimicrob Agents Chemother 49: 1865-1871. https://doi.org/10.1128/AAC.49.5.1865-1871.2005
|
| [47] |
Costa SS, Viveiros M, Amaral L, et al. (2013) Multidrug efflux pumps in Staphylococcus aureus: an update. Open Microbiol J 7: 59-71. https://doi.org/10.2174/1874285801307010059
|
| [48] |
Kaatz GW, McAleese F, Seo SM (2005) Multidrug resistance in Staphylococcus aureus due to overexpression of a novel multidrug and toxin extrusion (MATE) transport protein. Antimicrob Agents Chemother 49: 1857-1864. https://doi.org/10.1128/AAC.49.5.1857-1864.2005
|
| [49] |
Morita Y, Kataoka A, Shiota S, et al. (2000) NorM of Vibrio parahaemolyticus is an Na+-driven multidrug efflux pump. J Bacteriol 182: 6694-6697. https://doi.org/10.1128/JB.182.23.6694-6697.2000
|
| [50] |
Dridi L, Tankovic J, Petit JC (2004) CdeA of Clostridium difficile, a new multidrug efflux transporter of the MATE family. Microb Drug Resist 10: 191-196. https://doi.org/10.1089/mdr.2004.10.191
|
| [51] |
Kaatz GW, Moudgal VV, Seo SM, et al. (2003) Phenylpiperidine selective serotonin reuptake inhibitors interfere with multidrug efflux pump activity in Staphylococcus aureus. Int J Antimicrob Agents 22: 254-261. https://doi.org/10.1016/S0924-8579(03)00220-6
|
| [52] |
Schindler BD, Patel D, Seo SM, et al. (2013) Mutagenesis and modeling to predict structural and functional characteristics of the Staphylococcus aureus MepA multidrug efflux pump. J Bacteriol 195: 523-533. https://doi.org/10.1128/JB.01679-12
|
| [53] |
da Costa RHS, Rocha JE, de Freitas TS, et al. (2021) Evaluation of antibacterial activity and reversal of the NorA and MepA efflux pump of estragole against Staphylococcus aureus bacteria. Arch Microbiol 203: 3551-3555. https://doi.org/10.1007/s00203-021-02347-x
|
| [54] |
Oliveira-Tintino CDM, Tintino SR, Muniz DF, et al. (2021) Chemical synthesis, molecular docking and MepA efflux pump inhibitory effect by 1,8-naphthyridines sulfonamides. Eur J Pharm Sci 160: 105753. https://doi.org/10.1016/j.ejps.2021.105753
|
| [55] |
Freitas PR, de Araujo ACJ, Dos Santos Barbosa CR, et al. (2022) Inhibition of the MepA efflux pump by limonene demonstrated by in vitro and in silico methods. Folia Microbiol (Praha) 67: 15-20. https://doi.org/10.1007/s12223-021-00909-6
|
| [56] |
Wassenaar TM, Ussery D, Nielsen LN, et al. (2015) Review and phylogenetic analysis of qac genes that reduce susceptibility to quaternary ammonium compounds in Staphylococcus species. Eur J Microbiol Immunol (Bp) 5: 44-61. https://doi.org/10.1556/EuJMI-D-14-00038
|
| [57] |
Sasatsu M, Shima K, Shibata Y, et al. (1989) Nucleotide sequence of a gene that encodes resistance to ethidium bromide from a transferable plasmid in Staphylococcus aureus. Nucleic Acids Res 17: 10103. https://doi.org/10.1093/nar/17.23.10103
|
| [58] |
Grinius L, Dreguniene G, Goldberg EB, et al. (1992) A staphylococcal multidrug resistance gene product is a member of a new protein family. Plasmid 27: 119-129. https://doi.org/10.1016/0147-619X(92)90012-Y
|
| [59] |
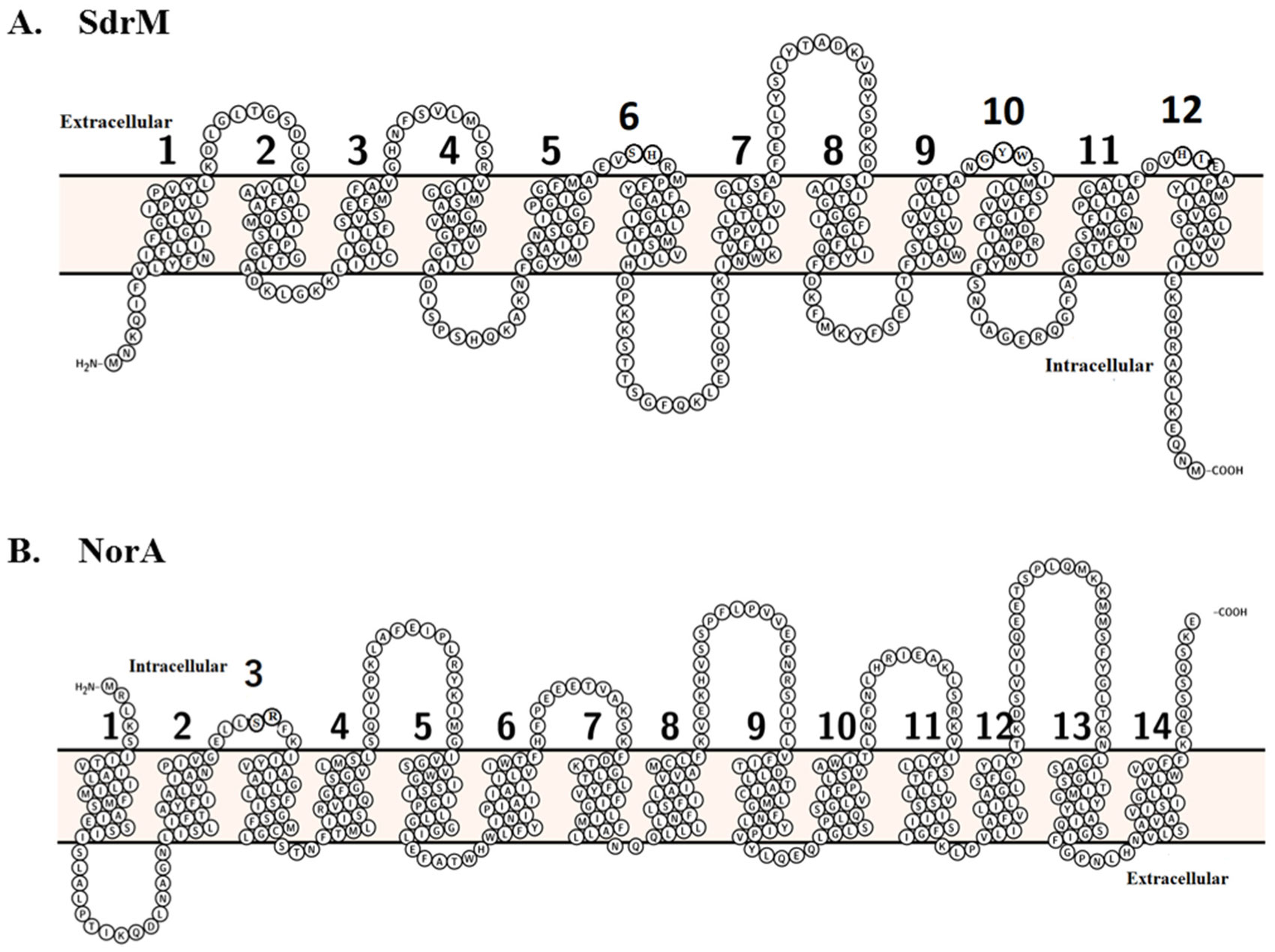
Yamada Y, Hideka K, Shiota S, et al. (2006) Gene cloning and characterization of SdrM, a chromosomally-encoded multidrug efflux pump, from Staphylococcus aureus. Biol Pharm Bull 29: 554-556. https://doi.org/10.1248/bpb.29.554
|
| [60] |
Grinius LL, Goldberg EB (1994) Bacterial multidrug resistance is due to a single membrane protein which functions as a drug pump. J Biol Chem 269: 29998-30004. https://doi.org/10.1016/S0021-9258(18)43980-4
|
| [61] |
Littlejohn TG, DiBerardino D, Messerotti LJ, et al. (1991) Structure and evolution of a family of genes encoding antiseptic and disinfectant resistance in Staphylococcus aureus. Gene 101: 59-66. https://doi.org/10.1016/0378-1119(91)90224-Y
|
| [62] |
Wassenaar TM, Ussery DW, Ingmer H (2016) The qacC gene has recently spread between rolling circle plasmids of Staphylococcus, indicative of a novel gene transfer mechanism. Front Microbiol 7: 1528. https://doi.org/10.3389/fmicb.2016.01528
|
| [63] |
Heir E, Sundheim G, Holck AL (1999) The qacG gene on plasmid pST94 confers resistance to quaternary ammonium compounds in staphylococci isolated from the food industry. J Appl Microbiol 86: 378-388. https://doi.org/10.1046/j.1365-2672.1999.00672.x
|
| [64] |
Heir E, Sundheim G, Holck AL (1998) The Staphylococcus qacH gene product: a new member of the SMR family encoding multidrug resistance. FEMS Microbiol Lett 163: 49-56. https://doi.org/10.1111/j.1574-6968.1998.tb13025.x
|
| [65] |
Jiang X, Xu Y, Li Y, et al. (2017) Characterization and horizontal transfer of qacH-associated class 1 integrons in Escherichia coli isolated from retail meats. Int J Food Microbiol 258: 12-17. https://doi.org/10.1016/j.ijfoodmicro.2017.07.009
|
| [66] |
Bjorland J, Steinum T, Sunde M, et al. (2003) Novel plasmid-borne gene qacJ mediates resistance to quaternary ammonium compounds in equine Staphylococcus aureus, Staphylococcus simulans, and Staphylococcus intermedius. Antimicrob Agents Chemother 47: 3046-3052. https://doi.org/10.1128/AAC.47.10.3046-3052.2003
|
| [67] |
Ninio S, Rotem D, Schuldiner S (2001) Functional analysis of novel multidrug transporters from human pathogens. J Biol Chem 276: 48250-48256. https://doi.org/10.1074/jbc.M108231200
|
| [68] |
Paulsen IT, Brown MH, Dunstan SJ, et al. (1995) Molecular characterization of the staphylococcal multidrug resistance export protein QacC. J Bacteriol 177: 2827-2833. https://doi.org/10.1128/jb.177.10.2827-2833.1995
|
| [69] |
Kumar S, Lekshmi M, Parvathi A, et al. (2020) Functional and structural roles of the major facilitator superfamily bacterial multidrug efflux pumps. Microorganisms 8: 266. https://doi.org/10.3390/microorganisms8020266
|
| [70] | Ranjana K, Shrestha U, Kumar S, et al. (2017) Molecular biology of multidrug resistance efflux pumps of the major facilitator superfamily from bacterial food pathogens. Foodborne Pathogens and Antibiotic Resistance . Wiley-Blackwell. https://doi.org/10.1002/9781119139188.ch13 |
| [71] |
Maiden MC, Davis EO, Baldwin SA, et al. (1987) Mammalian and bacterial sugar transport proteins are homologous. Nature 325: 641-643. https://doi.org/10.1038/325641a0
|
| [72] | Ranaweera I, Shrestha U, Ranjana KC, et al. (2015) Structural comparison of bacterial multidrug efflux pumps of the major facilitator superfamily. Trends Cell Mol Biol 10: 131-140. |
| [73] |
Henderson PJ (1990) The homologous glucose transport proteins of prokaryotes and eukaryotes. Res Microbiol 141: 316-328. https://doi.org/10.1016/0923-2508(90)90005-B
|
| [74] |
Griffith JK, Baker ME, Rouch DA, et al. (1992) Membrane transport proteins: implications of sequence comparisons. Curr Opin Cell Biol 4: 684-695. https://doi.org/10.1016/0955-0674(92)90090-Y
|
| [75] |
Henderson PJ (1991) Studies of translocation catalysis. Biosci Rep 11: 477-453. https://doi.org/10.1007/BF01130216
|
| [76] |
Henderson PJ (1990) Proton-linked sugar transport systems in bacteria. J Bioenerg Biomembr 22: 525-569. https://doi.org/10.1007/BF00762961
|
| [77] |
Varela MF, Griffith JK (1993) Nucleotide and deduced protein sequences of the class D tetracycline resistance determinant: Relationship to other antimicrobial transport proteins. Antimicrob Agents Chemother 37: 1253-1258. https://doi.org/10.1128/AAC.37.6.1253
|
| [78] |
Yoshida H, Bogaki M, Nakamura S, et al. (1990) Nucleotide sequence and characterization of the Staphylococcus aureus norA gene, which confers resistance to quinolones. J Bacteriol 172: 6942-6949. https://doi.org/10.1128/jb.172.12.6942-6949.1990
|
| [79] |
Omasits U, Ahrens CH, Muller S, et al. (2014) Protter: interactive protein feature visualization and integration with experimental proteomic data. Bioinformatics 30: 884-886. https://doi.org/10.1093/bioinformatics/btt607
|
| [80] |
Law CJ, Maloney PC, Wang DN (2008) Ins and outs of major facilitator superfamily antiporters. Annu Rev Microbiol 62: 289-305. https://doi.org/10.1146/annurev.micro.61.080706.093329
|
| [81] |
Yaffe D, Radestock S, Shuster Y, et al. (2013) Identification of molecular hinge points mediating alternating access in the vesicular monoamine transporter VMAT2. Proc Natl Acad Sci U S A 110: E1332-E1341. https://doi.org/10.1073/pnas.1220497110
|
| [82] |
Radestock S, Forrest LR (2011) The alternating-access mechanism of MFS transporters arises from inverted-topology repeats. J Mol Biol 407: 698-715. https://doi.org/10.1016/j.jmb.2011.02.008
|
| [83] | Kumar S, Ranjana KC, Sanford LM, et al. (2016) Structural and functional roles of two evolutionarily conserved amino acid sequence motifs within solute transporters of the major facilitator superfamily. Trends in Cell & Molecular Biology 11: 41-53. |
| [84] |
Truong-Bolduc QC, Hooper DC (2010) Phosphorylation of MgrA and its effect on expression of the NorA and NorB efflux pumps of Staphylococcus aureus. J Bacteriol 192: 2525-2534. https://doi.org/10.1128/JB.00018-10
|
| [85] |
Yamaguchi A, Shiina Y, Fujihira E, et al. (1995) The tetracycline efflux protein encoded by the tet(K) gene from Staphylococcus aureus is a metal-tetracycline/H+ antiporter. FEBS Lett 365: 193-197. https://doi.org/10.1016/0014-5793(95)00455-I
|
| [86] |
Jin J, Guffanti AA, Bechhofer DH, et al. (2002) Tet(L) and Tet(K) tetracycline-divalent metal/H+ antiporters: characterization of multiple catalytic modes and a mutagenesis approach to differences in their efflux substrate and coupling ion preferences. J Bacteriol 184: 4722-4732. https://doi.org/10.1128/JB.184.17.4722-4732.2002
|
| [87] |
Sheridan RP, Chopra I (1991) Origin of tetracycline efflux proteins: conclusions from nucleotide sequence analysis. Mol Microbiol 5: 895-900. https://doi.org/10.1111/j.1365-2958.1991.tb00763.x
|
| [88] |
Neyfakh AA, Borsch CM, Kaatz GW (1993) Fluoroquinolone resistance protein NorA of Staphylococcus aureus is a multidrug efflux transporter. Antimicrob Agents Chemother 37: 128-129. https://doi.org/10.1128/AAC.37.1.128
|
| [89] |
Neyfakh AA (1992) The multidrug efflux transporter of Bacillus subtilis is a structural and functional homolog of the Staphylococcus NorA protein. Antimicrob Agents Chemother 36: 484-485. https://doi.org/10.1128/AAC.36.2.484
|
| [90] |
Truong-Bolduc QC, Strahilevitz J, Hooper DC (2006) NorC, a new efflux pump regulated by MgrA of Staphylococcus aureus. Antimicrob Agents Chemother 50: 1104-1107. https://doi.org/10.1128/AAC.50.3.1104-1107.2006
|
| [91] |
Truong-Bolduc QC, Bolduc GR, Okumura R, et al. (2011) Implication of the NorB efflux pump in the adaptation of Staphylococcus aureus to growth at acid pH and in resistance to moxifloxacin. Antimicrob Agents Chemother 55: 3214-3219. https://doi.org/10.1128/AAC.00289-11
|
| [92] |
Ding Y, Fu Y, Lee JC, et al. (2012) Staphylococcus aureus NorD, a putative efflux pump coregulated with the Opp1 oligopeptide permease, contributes selectively to fitness in vivo. J Bacteriol 194: 6586-6593. https://doi.org/10.1128/JB.01414-12
|
| [93] | Brown MH, Skurray RA (2001) Staphylococcal multidrug efflux protein QacA. J Mol Microbiol Biotechnol 3: 163-170. |
| [94] |
Yamada Y, Shiota S, Mizushima T, et al. (2006) Functional gene cloning and characterization of MdeA, a multidrug efflux pump from Staphylococcus aureus. Biol Pharm Bull 29: 801-804. https://doi.org/10.1248/bpb.29.801
|
| [95] |
Huang J, O'Toole PW, Shen W, et al. (2004) Novel chromosomally encoded multidrug efflux transporter MdeA in Staphylococcus aureus. Antimicrob Agents Chemother 48: 909-917. https://doi.org/10.1128/AAC.48.3.909-917.2004
|
| [96] |
Floyd JL, Smith KP, Kumar SH, et al. (2010) LmrS is a multidrug efflux pump of the major facilitator superfamily from Staphylococcus aureus. Antimicrob Agents Chemother 54: 5406-5412. https://doi.org/10.1128/AAC.00580-10
|
| [97] | Truong-Bolduc QC, Wang Y, Hooper DC (2018) Tet38 efflux pump contributes to fosfomycin resistance in Staphylococcus aureus. Antimicrob Agents Chemother 62: e00927-18. https://doi.org/10.1128/AAC.00927-18 |
| [98] |
Majumder P, Khare S, Athreya A, et al. (2019) Dissection of protonation sites for antibacterial recognition and transport in QacA, a multi-drug efflux transporter. J Mol Biol 431: 2163-2179. https://doi.org/10.1016/j.jmb.2019.03.015
|
| [99] |
Kakarla P, Ranjana K, Shrestha U, et al. (2017) Functional roles of highly conserved amino acid sequence motifs A and C in solute transporters of the major facilitator superfamily. Drug resistance in bacteria, fungi, malaria, and cancer . Springer 111-140. https://doi.org/10.1007/978-3-319-48683-3_4
|
| [100] |
Shang Y, Lv P, Su D, et al. (2022) Evolutionary conservative analysis revealed novel functional sites in the efflux pump NorA of Staphylococcus aureus. J Antimicrob Chemother 77: 675-681. https://doi.org/10.1093/jac/dkab453
|
| [101] |
Varela MF, Sansom CE, Griffith JK (1995) Mutational analysis and molecular modelling of an amino acid sequence motif conserved in antiporters but not symporters in a transporter superfamily. Mol Membr Biol 12: 313-319. https://doi.org/10.3109/09687689509072433
|
| [102] |
Kakarla P, Floyd J, Mukherjee M, et al. (2017) Inhibition of the multidrug efflux pump LmrS from Staphylococcus aureus by cumin spice Cuminum cyminum. Arch Microbiol 199: 465-474. https://doi.org/10.1007/s00203-016-1314-5
|
| [103] | Truong-Bolduc QC, Wang Y, Chen C, et al. (2017) Transcriptional regulator TetR21 controls the expression of the Staphylococcus aureus LmrS efflux pump. Antimicrob Agents Chemother 61: e00649-17. https://doi.org/10.1128/AAC.00649-17 |
| [104] |
Nava AR, Mauricio N, Sanca AJ, et al. (2020) Evidence of calcium signaling and modulation of the LmrS multidrug resistant efflux pump activity by Ca2 + ions in S. aureus. Front Microbiol 11: 573388. https://doi.org/10.3389/fmicb.2020.573388
|
| [105] |
Pereira PS, Lima M, Neto PPM, et al. (2019) Thiazolidinedione and thiazole derivatives potentiate norfloxacin activity against NorA efflux pump over expression in Staphylococcus aureus 1199B strains. Bioorg Med Chem 27: 3797-3804. https://doi.org/10.1016/j.bmc.2019.07.006
|
| [106] |
Kalia NP, Mahajan P, Mehra R, et al. (2012) Capsaicin, a novel inhibitor of the NorA efflux pump, reduces the intracellular invasion of Staphylococcus aureus. J Antimicrob Chemother 67: 2401-2408. https://doi.org/10.1093/jac/dks232
|
| [107] |
Naaz F, Khan A, Kumari A, et al. (2021) 1,3,4-oxadiazole conjugates of capsaicin as potent NorA efflux pump inhibitors of Staphylococcus aureus. Bioorg Chem 113: 105031. https://doi.org/10.1016/j.bioorg.2021.105031
|
| [108] |
Zarate SG, Morales P, Swiderek K, et al. (2019) A molecular modeling approach to identify novel inhibitors of the major facilitator superfamily of efflux pump transporters. Antibiotics (Basel) 8: 25. https://doi.org/10.3390/antibiotics8010025
|
| [109] |
Shinnick SG, Varela MF (2002) Altered sugar selection and transport conferred by spontaneous point and deletion mutations in the lactose carrier of Escherichia coli. J Membr Biol 189: 191-199. https://doi.org/10.1007/s00232-002-1013-9
|
| [110] |
Varela MF, Wilson TH, Rodon-Rivera V, et al. (2000) Mutants of the lactose carrier of Escherichia coli which show altered sugar recognition plus a severe defect in sugar accumulation. J Membr Biol 174: 199-205. https://doi.org/10.1007/s002320001044
|
| [111] |
Varela MF, Brooker RJ, Wilson TH (1997) Lactose carrier mutants of Escherichia coli with changes in sugar recognition (lactose versus melibiose). J Bacteriol 179: 5570-5573. https://doi.org/10.1128/jb.179.17.5570-5573.1997
|
| [112] |
Muniz DF, Dos Santos Barbosa CR, de Menezes IRA, et al. (2021) In vitro and in silico inhibitory effects of synthetic and natural eugenol derivatives against the NorA efflux pump in Staphylococcus aureus. Food Chem 337: 127776. https://doi.org/10.1016/j.foodchem.2020.127776
|
| [113] |
Dos Santos Barbosa CR, Scherf JR, de Freitas TS, et al. (2021) Effect of Carvacrol and Thymol on NorA efflux pump inhibition in multidrug-resistant (MDR) Staphylococcus aureus strains. J Bioenerg Biomembr 53: 489-498. https://doi.org/10.1007/s10863-021-09906-3
|
| [114] |
Limaverde PW, Campina FF, da Cunha FAB, et al. (2017) Inhibition of the TetK efflux-pump by the essential oil of Chenopodium ambrosioides L. and α-terpinene against Staphylococcus aureus IS-58. Food Chem Toxicol 109: 957-961. https://doi.org/10.1016/j.fct.2017.02.031
|
| [115] |
Butaye P, Cloeckaert A, Schwarz S (2003) Mobile genes coding for efflux-mediated antimicrobial resistance in Gram-positive and Gram-negative bacteria. Int J Antimicrob Agents 22: 205-210. https://doi.org/10.1016/S0924-8579(03)00202-4
|
| [116] |
Piddock LJ (2006) Clinically relevant chromosomally encoded multidrug resistance efflux pumps in bacteria. Clin Microbiol Rev 19: 382-402. https://doi.org/10.1128/CMR.19.2.382-402.2006
|
| [117] |
Dos Santos JFS, Tintino SR, da Silva ARP, et al. (2021) Enhancement of the antibiotic activity by quercetin against Staphylococcus aureus efflux pumps. J Bioenerg Biomembr 53: 157-167. https://doi.org/10.1007/s10863-021-09886-4
|
| [118] |
Holler JG, Slotved HC, Molgaard P, et al. (2012) Chalcone inhibitors of the NorA efflux pump in Staphylococcus aureus whole cells and enriched everted membrane vesicles. Bioorg Med Chem 20: 4514-4521. https://doi.org/10.1016/j.bmc.2012.05.025
|
| [119] |
Rezende-Junior LM, Andrade LMS, Leal A, et al. (2020) Chalcones isolated from Arrabidaea brachypoda flowers as inhibitors of NorA and MepA multidrug efflux pumps of Staphylococcus aureus. Antibiotics (Basel) 9: 351. https://doi.org/10.3390/antibiotics9060351
|
| [120] |
Alves Borges Leal AL, Teixeira da Silva P, Nunes da Rocha M, et al. (2021) Potentiating activity of Norfloxacin by synthetic chalcones against NorA overproducing Staphylococcus aureus. Microb Pathog 155: 104894. https://doi.org/10.1016/j.micpath.2021.104894
|
| [121] |
Freitas TS, Xavier JC, Pereira RLS, et al. (2021) In vitro and in silico studies of chalcones derived from natural acetophenone inhibitors of NorA and MepA multidrug efflux pumps in Staphylococcus aureus. Microb Pathog 161: 105286. https://doi.org/10.1016/j.micpath.2021.105286
|
| [122] |
Rocha JE, de Freitas TS, da Cunha Xavier J, et al. (2021) Antibacterial and antibiotic modifying activity, ADMET study and molecular docking of synthetic chalcone (E)-1-(2-hydroxyphenyl)-3-(2,4-dimethoxy-3-methylphenyl)prop-2-en-1-one in strains of Staphylococcus aureus carrying NorA and MepA efflux pumps. Biomed Pharmacother 140: 111768. https://doi.org/10.1016/j.biopha.2021.111768
|
| [123] |
Oliveira-Tintino CDM, Muniz DF, Barbosa C, et al. (2021) The 1,8-naphthyridines sulfonamides are NorA efflux pump inhibitors. J Glob Antimicrob Resist 24: 233-240. https://doi.org/10.1016/j.jgar.2020.11.027
|
| [124] |
Pinheiro P, Santiago G, da Silva F, et al. (2021) Antibacterial activity and inhibition against Staphylococcus aureus NorA efflux pump by ferulic acid and its esterified derivatives. Asian Pac J Trop Biomed 11: 405-413. https://doi.org/10.4103/2221-1691.321130
|
| [125] |
Pinheiro PG, Santiago GMP, da Silva FEF, et al. (2022) Ferulic acid derivatives inhibiting Staphylococcus aureus TetK and MsrA efflux pumps. Biotechnol Rep 34: e00717. https://doi.org/10.1016/j.btre.2022.e00717
|
| [126] |
Figueredo FG, Ramos ITL, Paz JA, et al. (2020) Effect of hydroxyamines derived from lapachol and norlachol against Staphylococcus aureus strains carrying the NorA efflux pump. Infect Genet Evol 84: 104370. https://doi.org/10.1016/j.meegid.2020.104370
|
| [127] |
Pereira da Cruz R, Sampaio de Freitas T, Socorro Costa MD, et al. (2020) Effect of α-Bisabolol and its β-Cyclodextrin complex as TetK and NorA efflux pump inhibitors in Staphylococcus aureus strains. Antibiotics (Basel) 9: 28. https://doi.org/10.3390/antibiotics9010028
|
| [128] |
Rodrigues Dos Santos Barbosa C, Feitosa Muniz D, Silvino Pereira P, et al. (2021) Evaluation of Elaiophylin extracted from Streptomyces hygroscopicus as a potential inhibitor of the NorA efflux protein in Staphylococcus aureus: An in vitro and in silico approach. Bioorg Med Chem Lett 50: 128334. https://doi.org/10.1016/j.bmcl.2021.128334
|
| [129] |
Deka B, Suri M, Sarma S, et al. (2022) Potentiating the intracellular killing of Staphylococcus aureus by dihydroquinazoline analogues as NorA efflux pump inhibitor. Bioorg Med Chem 54: 116580. https://doi.org/10.1016/j.bmc.2021.116580
|
| [130] |
Holasová K, Křížkovská B, Hoang L, et al. (2022) Flavonolignans from silymarin modulate antibiotic resistance and virulence in Staphylococcus aureus. Biomed Pharmacother 149: 112806. https://doi.org/10.1016/j.biopha.2022.112806
|
| [131] |
Bruce SA, Smith JT, Mydosh JL, et al. (2022) Shared antibiotic resistance and virulence genes in Staphylococcus aureus from diverse animal hosts. Sci Rep 12: 4413. https://doi.org/10.1038/s41598-022-08230-z
|
| [132] | Truong-Bolduc QC, Wang Y, Hooper DC (2019) Tet38 of Staphylococcus aureus binds to host cell receptor complex CD36-Toll-Like receptor 2 and protects from teichoic acid synthesis inhibitors tunicamycin and congo red. Infect Immun 87: e00194-19. https://doi.org/10.1128/IAI.00194-19 |
| [133] |
Truong-Bolduc QC, Zhang X, Hooper DC (2003) Characterization of NorR protein, a multifunctional regulator of norA expression in Staphylococcus aureus. J Bacteriol 185: 3127-3138. https://doi.org/10.1128/JB.185.10.3127-3138.2003
|
| [134] |
Kumar S, He G, Kakarla P, et al. (2016) Bacterial multidrug efflux pumps of the major facilitator superfamily as targets for modulation. Infect Disord Drug Targets 16: 28-43. https://doi.org/10.2174/1871526516666160407113848
|
| [135] |
Kumar S, Varela MF (2012) Biochemistry of bacterial multidrug efflux pumps. Int J Mol Sci 13: 4484-4495. https://doi.org/10.3390/ijms13044484
|
| [136] |
Varela MF, Kumar S (2019) Strategies for discovery of new molecular targets for anti-infective drugs. Curr Opin Pharmacol 48: 57-68. https://doi.org/10.1016/j.coph.2019.04.015
|
| [137] |
Bame JR, Graf TN, Junio HA, et al. (2013) Sarothrin from Alkanna orientalis is an antimicrobial agent and efflux pump inhibitor. Planta Med 79: 327-329. https://doi.org/10.1055/s-0032-1328259
|
| [138] |
Buonerba F, Lepri S, Goracci L, et al. (2017) Improved potency of indole-based NorA efflux pump inhibitors: from serendipity toward rational design and development. J Med Chem 60: 517-523. https://doi.org/10.1021/acs.jmedchem.6b01281
|
| [139] |
Chan BC, Ip M, Lau CB, et al. (2011) Synergistic effects of baicalein with ciprofloxacin against NorA over-expressed methicillin-resistant Staphylococcus aureus (MRSA) and inhibition of MRSA pyruvate kinase. J Ethnopharmacol 137: 767-773. https://doi.org/10.1016/j.jep.2011.06.039
|
| [140] |
Coelho ML, Ferreira JH, de Siqueira Junior JP, et al. (2016) Inhibition of the NorA multi-drug transporter by oxygenated monoterpenes. Microb Pathog 99: 173-177. https://doi.org/10.1016/j.micpath.2016.08.026
|
| [141] |
Falcao-Silva VS, Silva DA, Souza Mde F, et al. (2009) Modulation of drug resistance in Staphylococcus aureus by a kaempferol glycoside from Herissantia tiubae (Malvaceae). Phytother Res 23: 1367-1370. https://doi.org/10.1002/ptr.2695
|
| [142] |
Hequet A, Burchak ON, Jeanty M, et al. (2014) 1-(1H-indol-3-yl)ethanamine derivatives as potent Staphylococcus aureus NorA efflux pump inhibitors. ChemMedChem 9: 1534-1545. https://doi.org/10.1002/cmdc.201400042
|
| [143] |
Holler JG, Christensen SB, Slotved HC, et al. (2012) Novel inhibitory activity of the Staphylococcus aureus NorA efflux pump by a kaempferol rhamnoside isolated from Persea lingue Nees. J Antimicrob Chemother 67: 1138-1144. https://doi.org/10.1093/jac/dks005
|
| [144] |
Kaatz GW, Seo SM, Ruble CA (1993) Efflux-mediated fluoroquinolone resistance in Staphylococcus aureus. Antimicrob Agents Chemother 37: 1086-1094. https://doi.org/10.1128/AAC.37.5.1086
|
| [145] |
Kumar A, Khan IA, Koul S, et al. (2008) Novel structural analogues of piperine as inhibitors of the NorA efflux pump of Staphylococcus aureus. J Antimicrob Chemother 61: 1270-1276. https://doi.org/10.1093/jac/dkn088
|
| [146] |
Lan JE, Li XJ, Zhu XF, et al. (2021) Flavonoids from Artemisia rupestris and their synergistic antibacterial effects on drug-resistant Staphylococcus aureus. Nat Prod Res 35: 1881-1886. https://doi.org/10.1080/14786419.2019.1639182
|
| [147] |
Lowrence RC, Raman T, Makala HV, et al. (2016) Dithiazole thione derivative as competitive NorA efflux pump inhibitor to curtail multi drug resistant clinical isolate of MRSA in a zebrafish infection model. Appl Microbiol Biotechnol 100: 9265-9281. https://doi.org/10.1007/s00253-016-7759-2
|
| [148] |
Mullin S, Mani N, Grossman TH (2004) Inhibition of antibiotic efflux in bacteria by the novel multidrug resistance inhibitors biricodar (VX-710) and timcodar (VX-853). Antimicrob Agents Chemother 48: 4171-4176. https://doi.org/10.1128/AAC.48.11.4171-4176.2004
|
| [149] |
Ponnusamy K, Ramasamy M, Savarimuthu I, et al. (2010) Indirubin potentiates ciprofloxacin activity in the NorA efflux pump of Staphylococcus aureus. Scand J Infect Dis 42: 500-505. https://doi.org/10.3109/00365541003713630
|
| [150] |
Randhawa HK, Hundal KK, Ahirrao PN, et al. (2016) Efflux pump inhibitory activity of flavonoids isolated from Alpinia calcarata against methicillin-resistant Staphylococcus aureus. Biologia 71: 484-493. https://doi.org/10.1515/biolog-2016-0073
|
| [151] |
Braga Ribeiro AM, Sousa JN, Costa LM, et al. (2019) Antimicrobial activity of Phyllanthus amarus Schumach. & Thonn and inhibition of the NorA efflux pump of Staphylococcus aureus by Phyllanthin. Microb Pathog 130: 242-246. https://doi.org/10.1016/j.micpath.2019.03.012
|
| [152] |
Roy SK, Kumari N, Pahwa S, et al. (2013) NorA efflux pump inhibitory activity of coumarins from Mesua ferrea. Fitoterapia 90: 140-150. https://doi.org/10.1016/j.fitote.2013.07.015
|
| [153] |
Sabatini S, Gosetto F, Manfroni G, et al. (2011) Evolution from a natural flavones nucleus to obtain 2-(4-Propoxyphenyl)quinoline derivatives as potent inhibitors of the S. aureus NorA efflux pump. J Med Chem 54: 5722-5736. https://doi.org/10.1021/jm200370y
|
| [154] |
Samosorn S, Bremner JB, Ball A, et al. (2006) Synthesis of functionalized 2-aryl-5-nitro-1H-indoles and their activity as bacterial NorA efflux pump inhibitors. Bioorg Med Chem 14: 857-865. https://doi.org/10.1016/j.bmc.2005.09.019
|
| [155] |
Shiu WK, Malkinson JP, Rahman MM, et al. (2013) A new plant-derived antibacterial is an inhibitor of efflux pumps in Staphylococcus aureus. Int J Antimicrob Agents 42: 513-518. https://doi.org/10.1016/j.ijantimicag.2013.08.007
|
| [156] |
Smith EC, Kaatz GW, Seo SM, et al. (2007) The phenolic diterpene totarol inhibits multidrug efflux pump activity in Staphylococcus aureus. Antimicrob Agents Chemother 51: 4480-4483. https://doi.org/10.1128/AAC.00216-07
|
| [157] |
Stermitz FR, Scriven LN, Tegos G, et al. (2002) Two flavonols from Artemisa annua which potentiate the activity of berberine and norfloxacin against a resistant strain of Staphylococcus aureus. Planta Med 68: 1140-1141. https://doi.org/10.1055/s-2002-36347
|
| [158] |
Tintino SR, Oliveira-Tintino CD, Campina FF, et al. (2016) Evaluation of the tannic acid inhibitory effect against the NorA efflux pump of Staphylococcus aureus. Microb Pathog 97: 9-13. https://doi.org/10.1016/j.micpath.2016.04.003
|
| [159] |
Tintino SR, Souza VCA, Silva J, et al. (2020) Effect of Vitamin K3 inhibiting the function of NorA efflux pump and its gene expression on Staphylococcus aureus. Membranes (Basel) 10: 130. https://doi.org/10.3390/membranes10060130
|
| [160] |
Waditzer M, Bucar F (2021) Flavonoids as inhibitors of bacterial efflux pumps. Molecules 26: 6904. https://doi.org/10.3390/molecules26226904
|
| [161] | Wang D, Xie K, Zou D, et al. (2018) Inhibitory effects of silybin on the efflux pump of methicillin resistant Staphylococcus aureus. Mol Med Rep 18: 827-833. https://doi.org/10.3892/mmr.2018.9021 |
| [162] |
Wani NA, Singh S, Farooq S, et al. (2016) Amino acid amides of piperic acid (PA) and 4-ethylpiperic acid (EPA) as NorA efflux pump inhibitors of Staphylococcus aureus. Bioorg Med Chem Lett 26: 4174-4178. https://doi.org/10.1016/j.bmcl.2016.07.062
|
| [163] |
Yu JL, Grinius L, Hooper DC (2002) NorA functions as a multidrug efflux protein in both cytoplasmic membrane vesicles and reconstituted proteoliposomes. J Bacteriol 184: 1370-1377. https://doi.org/10.1128/JB.184.5.1370-1377.2002
|
| [164] |
Zhang J, Sun Y, Wang Y, et al. (2014) Non-antibiotic agent ginsenoside 20(S)-Rh2 enhanced the antibacterial effects of ciprofloxacin in vitro and in vivo as a potential NorA inhibitor. Eur J Pharmacol 740: 277-284. https://doi.org/10.1016/j.ejphar.2014.07.020
|
| [165] |
Joshi P, Singh S, Wani A, et al. (2014) Osthol and curcumin as inhibitors of human Pgp and multidrug efflux pumps of Staphylococcus aureus: reversing the resistance against frontline antibacterial drugs. MedChemComm 5: 1540-1547. https://doi.org/10.1039/C4MD00196F
|
| [166] |
Shokoofeh N, Moradi-Shoeili Z, Naeemi AS, et al. (2019) Biosynthesis of Fe3O4@Ag nanocomposite and evaluation of its performance on expression of norA and norB efflux pump genes in ciprofloxacin-resistant Staphylococcus aureus. Biol Trace Elem Res 191: 522-530. https://doi.org/10.1007/s12011-019-1632-y
|
| [167] |
Gupta VK, Tiwari N, Gupta P, et al. (2016) A clerodane diterpene from Polyalthia longifolia as a modifying agent of the resistance of methicillin resistant Staphylococcus aureus. Phytomedicine 23: 654-661. https://doi.org/10.1016/j.phymed.2016.03.001
|
| [168] |
Truong-Bolduc QC, Bolduc GR, Medeiros H, et al. (2015) Role of the Tet38 efflux pump in Staphylococcus aureus internalization and survival in epithelial cells. Infect Immun 83: 4362-4372. https://doi.org/10.1128/IAI.00723-15
|
| [169] |
Kumar S, Athreya A, Gulati A, et al. (2021) Structural basis of inhibition of a transporter from Staphylococcus aureus, NorC, through a single-domain camelid antibody. Commun Biol 4: 836. https://doi.org/10.1038/s42003-021-02357-x
|
| [170] |
Jang S (2016) Multidrug efflux pumps in Staphylococcus aureus and their clinical implications. J Microbiol 54: 1-8. https://doi.org/10.1007/s12275-016-5159-z
|
| [171] |
Mirza ZM, Kumar A, Kalia NP, et al. (2011) Piperine as an inhibitor of the MdeA efflux pump of Staphylococcus aureus. J Med Microbiol 60: 1472-1478. https://doi.org/10.1099/jmm.0.033167-0
|
| [172] |
Rouch DA, Cram DS, DiBerardino D, et al. (1990) Efflux-mediated antiseptic resistance gene qacA from Staphylococcus aureus: common ancestry with tetracycline- and sugar-transport proteins. Mol Microbiol 4: 2051-2062. https://doi.org/10.1111/j.1365-2958.1990.tb00565.x
|
| [173] |
Littlejohn TG, Paulsen IT, Gillespie MT, et al. (1992) Substrate specificity and energetics of antiseptic and disinfectant resistance in Staphylococcus aureus. FEMS Microbiol Lett 74: 259-265. https://doi.org/10.1111/j.1574-6968.1992.tb05376.x
|
| [174] | Tennent JM, Lyon BR, Midgley M, et al. (1989) Physical and biochemical characterization of the qacA gene encoding antiseptic and disinfectant resistance in Staphylococcus aureus. J Gen Microbiol 135: 1-10. https://doi.org/10.1099/00221287-135-1-1 |
| [175] |
AKF ES, Dos Reis AC, Pinheiro EEA, et al. (2021) Modulation of the drug resistance by Platonia insignis Mart. Extract, ethyl acetate fraction and morelloflavone/volkensiflavone (biflavonoids) in Staphylococcus aureus strains overexpressing efflux pump genes. Curr Drug Metab 22: 114-122. https://doi.org/10.2174/1389200221666200523155617
|
| [176] |
Mitchell BA, Paulsen IT, Brown MH, et al. (1999) Bioenergetics of the staphylococcal multidrug export protein QacA. Identification of distinct binding sites for monovalent and divalent cations. J Biol Chem 274: 3541-3548. https://doi.org/10.1074/jbc.274.6.3541
|
| [177] | Dymek A, Armada A, Handzlik J, et al. (2012) The activity of 16 new hydantoin compounds on the intrinsic and overexpressed efflux pump system of Staphylococcus aureus. In Vivo 26: 223-229. |
| [178] |
Truong-Bolduc QC, Dunman PM, Strahilevitz J, et al. (2005) MgrA is a multiple regulator of two new efflux pumps in Staphylococcus aureus. J Bacteriol 187: 2395-2405. https://doi.org/10.1128/JB.187.7.2395-2405.2005
|
| [179] |
Guay GG, Tuckman M, McNicholas P, et al. (1993) The tet(K) gene from Staphylococcus aureus mediates the transport of potassium in Escherichia coli. J Bacteriol 175: 4927-4929. https://doi.org/10.1128/jb.175.15.4927-4929.1993
|
| [180] |
Ginn SL, Brown MH, Skurray RA (2000) The TetA(K) tetracycline/H+ antiporter from Staphylococcus aureus: mutagenesis and functional analysis of motif C. J Bacteriol 182: 1492-1498. https://doi.org/10.1128/JB.182.6.1492-1498.2000
|
| [181] |
Gibbons S, Oluwatuyi M, Veitch NC, et al. (2003) Bacterial resistance modifying agents from Lycopus europaeus. Phytochemistry 62: 83-87. https://doi.org/10.1016/S0031-9422(02)00446-6
|
| [182] |
Zhu Y, Wang C, Schwarz S, et al. (2021) Identification of a novel tetracycline resistance gene, tet(63), located on a multiresistance plasmid from Staphylococcus aureus. J Antimicrob Chemother 76: 576-581. https://doi.org/10.1093/jac/dkaa485
|
| [183] |
Kehrenberg C, Schwarz S (2004) fexA, a novel Staphylococcus lentus gene encoding resistance to florfenicol and chloramphenicol. Antimicrob Agents Chemother 48: 615-618. https://doi.org/10.1128/AAC.48.2.615-618.2004
|
| [184] |
Mancuso G, Midiri A, Gerace E, et al. (2021) Bacterial antibiotic resistance: the most critical pathogens. Pathogens 10: 1310. https://doi.org/10.3390/pathogens10101310
|
| [185] |
Lekshmi M, Ammini P, Kumar S, et al. (2017) The food production environment and the development of antimicrobial resistance in human pathogens of animal origin. Microorganisms 5: 11. https://doi.org/10.3390/microorganisms5010011
|
| [186] | Varela MF, Andersen JL, Ranjana K, et al. (2017) Bacterial resistance mechanisms and inhibitors of multidrug efflux pumps belonging to the major facilitator superfamily of solute transport systems. Frontiers in Anti-Infective Drug Discovery . Bentham Science Publishers 109-131. https://doi.org/10.2174/9781681082912117050006 |
| [187] | Floyd JT, Kumar S, Mukherjee MM, et al. (2013) A review of the molecular mechanisms of drug efflux in pathogenic bacteria: A structure-function perspective. Recent Research Developments in Membrane Biology . Research Signpost, Inc. 15-66. |
| [188] |
Mukherjee M, Kakarla P, Kumar S, et al. (2014) Comparative genome analysis of non-toxigenic non-O1 versus toxigenic O1 Vibrio cholerae. Genom Discov 2: 1-15. https://doi.org/10.7243/2052-7993-2-1
|
| [189] | Kumar S, Lindquist IE, Sundararajan A, et al. (2013) Genome Sequence of Non-O1 Vibrio cholerae PS15. Genome Announc 1: e00227-12. https://doi.org/10.1128/genomeA.00227-12 |
| [190] |
Kumar S, Smith KP, Floyd JL, et al. (2011) Cloning and molecular analysis of a mannitol operon of phosphoenolpyruvate-dependent phosphotransferase (PTS) type from Vibrio cholerae O395. Arch Microbiol 193: 201-208. https://doi.org/10.1007/s00203-010-0663-8
|
| [191] |
Varela MF (2019) Antimicrobial efflux pumps. Antibiotic Drug Resistance . John Wiley & Sons, Inc. 167-179. https://doi.org/10.1002/9781119282549.ch8
|
| [192] | Rao M, Padyana S, Dipin K, et al. (2018) Antimicrobial compounds of plant origin as efflux pump inhibitors: new avenues for controlling multidrug resistant pathogens. J Antimicrob Agents 4: 1000159. https://doi.org/10.4172/2472-1212.1000159 |
| [193] | Shrestha U, Lekshmi M, Kumar S, et al. (2018) Bioactive agents as modulators of multidrug efflux pumps of the major facilitator superfamily in key bacterial pathogens. Curr Trends Microbiol 12: 15-37. |
| [194] |
Stephen J, Lekshmi M, Ammini P, et al. (2022) Membrane efflux pumps of pathogenic Vibrio species: role in antimicrobial resistance and virulence. Microorganisms 10: 382. https://doi.org/10.3390/microorganisms10020382
|
| [195] | Stephen J, Mukherjee S, Lekshmi M, et al. (2020) Antibiotic resistance in fish-borne pathogens of public health significance: An emerging food safety issue. Trends Microbiol 14: 11-20. |
| [196] |
Kumar S, Mukherjee MM, Varela MF (2013) Modulation of bacterial multidrug resistance efflux pumps of the major facilitator superfamily. Int J Bacteriol 2013: 204141. https://doi.org/10.1155/2013/204141
|
Figures(3) / Tables(1)

Manjusha Lekshmi, Jerusha Stephen, Manisha Ojha, Sanath Kumar, Manuel Varela. Staphylococcus aureus antimicrobial efflux pumps and their inhibitors: recent developments[J]. AIMS Medical Science, 2022, 9(3): 367-393. doi: 10.3934/medsci.2022018







 DownLoad:
DownLoad: