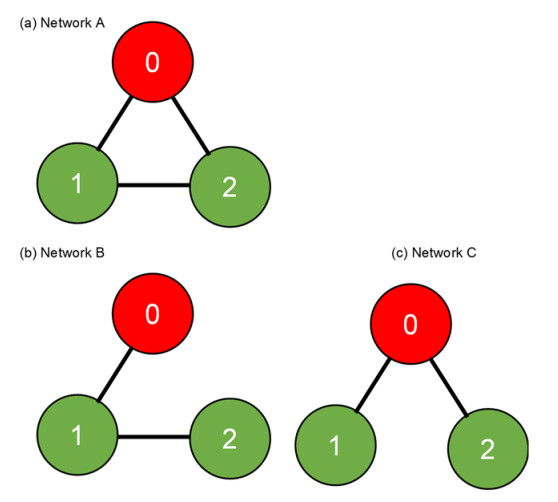
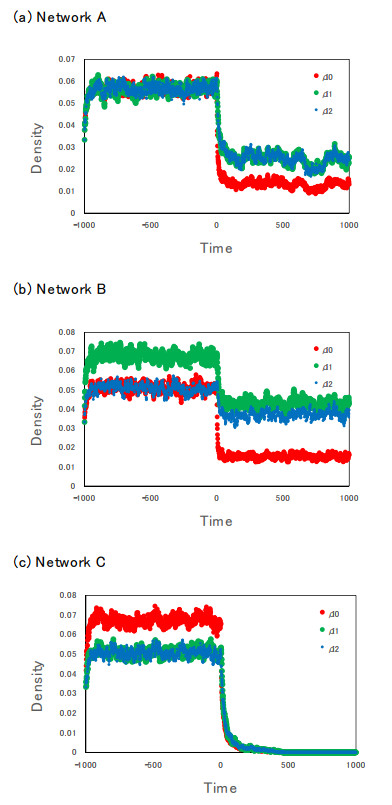
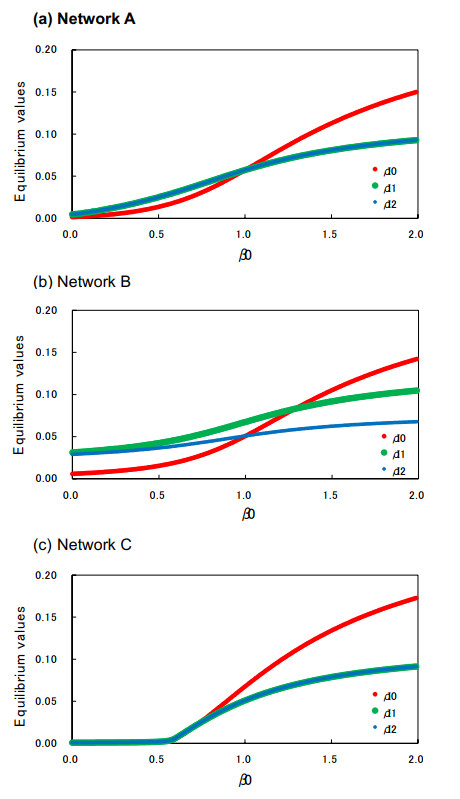
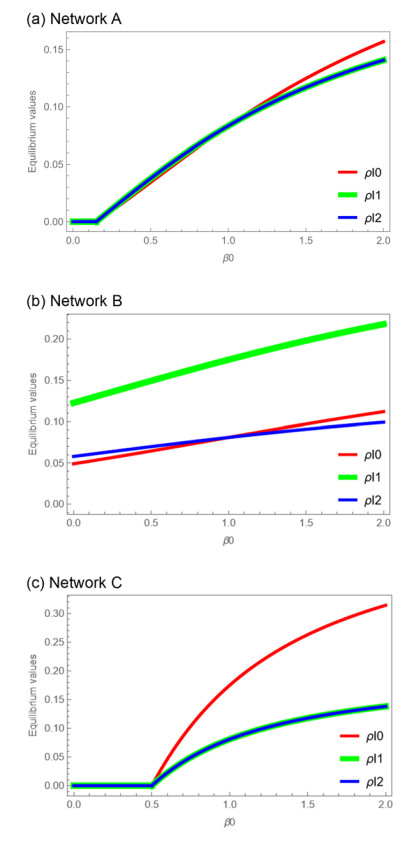
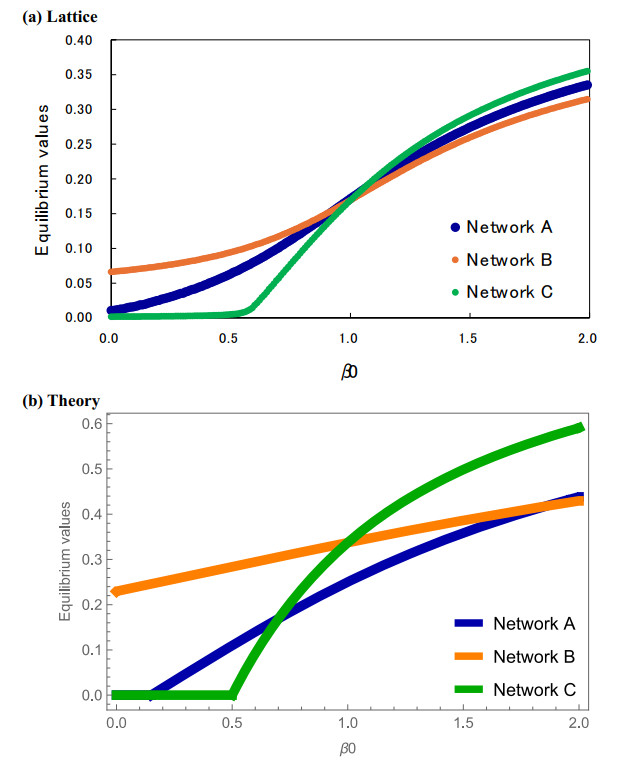
Control of infectious disease is very hard but important for the life of human beings. We study the susceptible–infected–susceptible (SIS) model of three-city networks. The SIS model simply contains both infection and recovery processes. We assume that human beings ("agents") live in three spatially separated cities, and they randomly migrate between cities. Two methods are applied: one is a computer simulation of an agent-based model, and the other is the theory of metapopulation dynamics. Both the simulation and theory reveal that the "hub city" plays an important role for disease control. It was found that we can eliminate the entire infection by disease control measures on the hub city only. Moreover, we found a paradoxical result: increased agent interaction does not necessarily lead to the spread of infection.
Citation: Nariyuki Nakagiri, Hiroki Yokoi, Yukio Sakisaka, Kei-ichi Tainaka, Kazunori Sato. Influence of network structure on infectious disease control[J]. Mathematical Biosciences and Engineering, 2025, 22(4): 943-961. doi: 10.3934/mbe.2025034
Control of infectious disease is very hard but important for the life of human beings. We study the susceptible–infected–susceptible (SIS) model of three-city networks. The SIS model simply contains both infection and recovery processes. We assume that human beings ("agents") live in three spatially separated cities, and they randomly migrate between cities. Two methods are applied: one is a computer simulation of an agent-based model, and the other is the theory of metapopulation dynamics. Both the simulation and theory reveal that the "hub city" plays an important role for disease control. It was found that we can eliminate the entire infection by disease control measures on the hub city only. Moreover, we found a paradoxical result: increased agent interaction does not necessarily lead to the spread of infection.

| [1] | R. M. Anderson, R. M. May, Infectious Diseases of Humans, Oxford University Press, Oxford, 1991. |
| [2] |
L. Wang, J. T. Wu, Characterizing the dynamics underlying global spread of epidemics, Nat. Commun., 9 (2018), 218. https://doi.org/10.1038/s41467-017-02344-z doi: 10.1038/s41467-017-02344-z

|
| [3] |
F. Ball, D. Sirl, Evaluation of vaccination strategies for SIR epidemics on random networks incorporating household structure, J. Math. Biol., 76 (2018), 483–530. https://doi.org/10.1007/s00285-017-1139-0 doi: 10.1007/s00285-017-1139-0
|
| [4] |
H. W. Hethcote, The mathematics of infectious diseases, SIAM Rev., 42 (2000), 599–653. https://doi.org/10.1137/S0036144500371907 doi: 10.1137/S0036144500371907
|
| [5] | J. D. Murray, Mathematical Biology, Springer-Verlag, 2002. https://doi.org/10.1007/b98868 |
| [6] |
N. Sharma, A. K. Gupta, Impact of time delay on the dynamics of SEIR epidemic model using cellular automata, Physica A, 471 (2017), 114–125. https://doi.org/10.1016/j.physa.2016.12.010 doi: 10.1016/j.physa.2016.12.010
|
| [7] |
A. A. Khan, R. Amin, S. Ullah, W. Sumelka, M. Altanji, Numerical simulation of a Caputo fractional epidemic model for the novel corona virus with the impact of environmental transmission, Alexandria Eng. J., 61 (2022), 5083–5095. https://doi.org/10.1016/j.aej.2021.10.008 doi: 10.1016/j.aej.2021.10.008
|
| [8] |
N. Nakagiri, H. Yokoi, A. Morishita, K. Tainaka, Three-lattice metapopulation model: Connecting corridor between patches may be harmful due to "hub effect", Ecol. Complexity, 59 (2024), 101090. https://doi.org/10.1016/j.ecocom.2024.101090 doi: 10.1016/j.ecocom.2024.101090
|
| [9] |
T. E. Harris, Contact interactions on a lattice, Ann. Probab., 2 (1974), 969–988. https://doi.org/10.1214/aop/1176996493 doi: 10.1214/aop/1176996493
|
| [10] |
Y. Itoh, K. Tainaka, T. Sakata, T. Tao, N. Nakagiri, Spatial enhancement of population uncertainty near the extinction threshold, Ecol. Modell., 174 (2004), 191–201. https://doi.org/10.1016/j.ecolmodel.2004.01.004 doi: 10.1016/j.ecolmodel.2004.01.004
|
| [11] |
A. Viguerie, G. Lorenzo, F. Auricchio, D. Baroli, T. J. R. Hughes, A. Patton, et al., Simulating the spread of COVID-19 via a spatially-resolved susceptible-exposed-infected-recovered-deceased (SEIRD) model with heterogeneous diffusion, Appl. Math. Lett., 111 (2021), 106617. https://doi.org/10.1016/j.aml.2020.106617 doi: 10.1016/j.aml.2020.106617
|
| [12] |
G. Bertaglia, L. Pareschi, Hyperbolic models for the spread of epidemics on networks: kinetic description and numerical methods, ESAIM Math. Model. Numer. Anal., 55 (2021), 381–407. https://doi.org/10.1051/m2an/2020082 doi: 10.1051/m2an/2020082
|
| [13] |
G. Bertaglia, L. Pareschi, Hyperbolic compartmental models for epidemic spread on networks with uncertain data: application to the emergence of COVID-19 in Italy, Math. Models Methods Appl. Sci., 31 (2021), 2495–2531. https://doi.org/10.1142/S0218202521500548 doi: 10.1142/S0218202521500548
|
| [14] |
G. Dimarco, B. Perthame, G. Toscani, M. Zanella, Kinetic models for epidemic dynamics with social heterogeneity, J. Math. Biol., 83 (2021), 4. https://doi.org/10.1007/s00285-021-01630-1 doi: 10.1007/s00285-021-01630-1
|
| [15] | G. Albi, G. Bertaglia, W. Boscheri, G. Dimarco, L. Pareschi, G. Toscani, et al., Kinetic modelling of epidemic dynamics: social contacts, control with uncertain data, and multiscale spatial dynamics, in Predicting Pandemics in a Globally Connected World (eds. N. Bellomo, M. A. J. Chaplain), Springer Nature, (2022), 43–108. https://doi.org/10.1007/978-3-030-96562-4_3 |
| [16] | I. Hanski, O. E. Gaggiotti, Ecology, Genetics, and Evolution of Metapopulations, Burlington, MA: Elsevier, 2004. https://doi.org/10.1016/B978-012323448-3/50003-9 |
| [17] | I. Hanski, M. E. Gilpin, Metapopulation Biology: Ecology, Genetics, and Evolution, San Diego, CA: Academic Press, 1997. |
| [18] |
S. A. Levin, Dispersion and population interactions, Am. Nat., 108 (1974), 207–228. https://doi.org/10.1086/282900 doi: 10.1086/282900
|
| [19] |
G. Cantin, N. Verdière, Networks of forest ecosystems: Mathematical modeling of their biotic pump mechanism and resilience to certain patch deforestation, Ecol. Complexity, 43 (2020), 100850. https://doi.org/10.1016/j.ecocom.2020.100850 doi: 10.1016/j.ecocom.2020.100850
|
| [20] |
N. Nakagiri, H. Yokoi, Y. Sakisaka, K. Tainaka, Population persistence under two conservation measures: paradox of habitat protection in patchy environment, Math. Biosci. Eng., 19 (2022), 9244–9257. https://doi.org/10.3934/mbe.2022429 doi: 10.3934/mbe.2022429
|
| [21] |
N. Masuda, M. A. Porter, R. Lambiotte, Random walks and diffusion on networks, Phys. Rep. 716–717 (2017), 1–58. https://doi.org/10.1016/j.physrep.2017.07.007 doi: 10.1016/j.physrep.2017.07.007
|
| [22] |
T. Nagatani, G. Ichinose, K. Tainaka, Epidemics of random walkers in metapopulation model for complete, cycle, and star graphs, J. Theor. Biol., 450 (2018), 66–75. https://doi.org/10.1016/j.jtbi.2018.04.029 doi: 10.1016/j.jtbi.2018.04.029
|
| [23] |
T. Nagatani, G. Ichinose, K. Tainaka, Heterogeneous network promotes species coexistence: metapopulation model for rock-paper-scissors game, Sci. Rep., 8 (2018), 7094. https://doi.org/10.1038/s41598-018-25353-4 doi: 10.1038/s41598-018-25353-4
|
| [24] |
H. Yokoi, K. Tainaka, K. Sato, Metapopulation model for a prey-predator system: Nonlinear migration due to the finite capacities of patches, J. Theor. Biol., 477 (2019), 24–35. https://doi.org/10.1016/j.jtbi.2019.05.021 doi: 10.1016/j.jtbi.2019.05.021
|
| [25] |
K. Yokoi, K. Tainaka, N. Nakagiri, K. Sato, Self-organized habitat segregation in an ambush-predator system: nonlinear migration of prey between two patches with finite capacities, Ecol. Inf., 55 (2020), 101022. https://doi.org/10.1016/j.ecoinf.2021.101477 doi: 10.1016/j.ecoinf.2021.101477
|
| [26] |
J. Menezes, S. Rodrigues, S. Batista, Mobility unevenness in rock–paper–scissors models, Ecol. Complexity, 52 (2022) 101028. https://doi.org/10.1016/j.ecocom.2022.101028 doi: 10.1016/j.ecocom.2022.101028
|
| [27] |
A. Mohammadi, K. Almasieh, D. Nayeri, F. Ataei, A. Khani, J. V. López-Bao, et al., Identifying priority core habitats and corridors for effective conservation of brown bears in Iran, Sci. Rep., 11 (2021), 1044. https://doi.org/10.1038/s41598-020-79970-z doi: 10.1038/s41598-020-79970-z
|
| [28] |
E. Jacob, P. Mörters, The contact process on scale-free networks evolving by vertex updating, R. Soc. Open Sci., 4 (2017), 170081. https://doi.org/10.1098/rsos.170081 doi: 10.1098/rsos.170081
|
| [29] |
R. Juhász, F. Iglói, Mixed-order phase transition of the contact process near multiple junctions, Phys. Rev. E, 95 (2017), 022109. https://doi.org/10.1103/PhysRevE.95.022109 doi: 10.1103/PhysRevE.95.022109
|
| [30] |
M. Katori, N. Konno, Upper bounds for survival probability of the contact process, J. Stat. Phys., 63 (1991), 115–130. https://doi.org/10.1007/BF01026595 doi: 10.1007/BF01026595
|
| [31] | J. Marro, R. Dickman, Nonequilibrium Phase Transition in Lattice Models, Cambridge University Press, Cambridge, 1999. |
| [32] |
C. Pu, S. Li, J. Yang, Epidemic spreading driven by biased random walks, Physica A, 432 (2015), 230–239. https://doi.org/10.1016/j.physa.2015.03.035 doi: 10.1016/j.physa.2015.03.035
|
| [33] |
Q. Zhang, K. Sun, M. Chinazzi, A. Pastore y Piontti, N. E. Dean, D. P. Rojas, et al., Spread of Zika virus in the Americas, Proc. Natl. Acad. Sci., 114 (2017), E4334–E4343. https://doi.org/10.1073/pnas.1620161114 doi: 10.1073/pnas.1620161114
|
| [34] |
S. Allesina, J. M. Levine, A competitive network theory of species diversity, Proc. Natl. Acad. Sci., 108 (2011), 56385642. https://doi.org/10.1073/pnas.1108946108 doi: 10.1073/pnas.1108946108
|
| [35] |
M. Kivelä, A. Arenas, M. Barthelemy, J. P. Gleeson, Y. Moreno, M. A. Porter, Multilayer networks, J. Complex Networks, 2 (2014), 203–271. https://doi.org/10.1093/comnet/cnu016 doi: 10.1093/comnet/cnu016
|
| [36] | A. L. Barabási, M. Pósfai, Network Science, Cambridge University Press, Cambridge, 2016. |
| [37] |
T. L. Czárán, R. F. Hoekstra, Killer-sensitive coexistence in metapopulations of micro-organisms, Proc. R. Soc. Lond. B, 270 (2003), 1373–1378. https://doi.org/10.1098/rspb.2003.2338 doi: 10.1098/rspb.2003.2338
|
| [38] |
T. Nagatani, G. Ichinose, Diffusively-coupled rock-paper-scissors game with mutation in scale-free hierarchical networks, Complexity, (2020), 6976328. https://doi.org/10.1155/2020/6976328 doi: 10.1155/2020/6976328
|
| [39] |
K. Tainaka, N. Nakagiri, H. Yokoi, K. Sato, Multi-layered model for rock-paper-scissors game: a swarm intelligence sustains biodiversity, Ecol. Inf., 66 (2021), 101477. https://doi.org/10.1016/j.ecoinf.2021.101477 doi: 10.1016/j.ecoinf.2021.101477
|
| [40] |
K. M. A. Kabir, J. Tanimoto, Impact of awareness in metapopulation epidemic model to suppress the infected individuals for different graphs, Eur. Phys. J. B, 92 (2019), 199. https://doi.org/10.1140/epjb/e2019-90570-7 doi: 10.1140/epjb/e2019-90570-7
|
| [41] |
K. M. A. Kabir, J. Tanimoto, Evolutionary vaccination game approach in metapopulation migration model with information spreading on different graphs, Chaos, Solitons Fractals, 120 (2019), 41–55. https://doi.org/10.1016/j.chaos.2019.01.013 doi: 10.1016/j.chaos.2019.01.013
|
| [42] |
C. Hui, M. A. McGeoch, Spatial patterns of Prisoner's dilemma game in metapopulation, Bull. Math. Biol., 69 (2007), 659–676. https://doi.org/10.1007/s11538-006-9145-1 doi: 10.1007/s11538-006-9145-1
|
| [43] |
K. Tainaka, Lattice model for the Lotka-Volterra system, J. Phys. Soc. Jpn., 57 (1988), 2588–2590. https://doi.org/10.1143/JPSJ.57.2588 doi: 10.1143/JPSJ.57.2588
|
| [44] |
D. L. Deangelis, W. M. Mooij, Individual-based modeling of ecological and evolutionary processes, Ann. Rev. Ecol. Evol. Syst., 36 (2005), 147–168. https://doi.org/10.1146/annurev.ecolsys.36.102003.152644 doi: 10.1146/annurev.ecolsys.36.102003.152644
|
| [45] |
A. R. Mikler, S. Venkatachalam, K. Abbas, modeling infectious diseases using global stochastic cellular automata, J. Biol. Syst., 13 (2005), 421–439. https://doi.org/10.1142/S0218339005001604 doi: 10.1142/S0218339005001604
|
| [46] |
A. K. Gupta, P. Redhu, Analyses of driver's anticipation effect in sensing relative flux in a new lattice model for two-lane traffic system, Physica A, 392 (2013), 5622–5632. https://doi.org/10.1016/j.physa.2013.07.040 doi: 10.1016/j.physa.2013.07.040
|
| [47] |
M. Perc, A. Szolnoki, Coevolutionary games—A mini review, Biosystems, 99 (2010), 109–125. https://doi.org/10.1016/j.biosystems.2009.10.003 doi: 10.1016/j.biosystems.2009.10.003
|
| [48] |
R. C. Alamino, An agent-based lattice model for the emergence of anti-microbial resistance, J. Theor. Biol., 486 (2019), 110080. https://doi.org/10.1016/j.jtbi.2019.110080 doi: 10.1016/j.jtbi.2019.110080
|
| [49] |
D. Miyagawa, G. Ichinose, Cellular automaton model with turning behavior in crowd evacuation, Physica A, 549 (2020) 124376. https://doi.org/10.1016/j.physa.2020.124376 doi: 10.1016/j.physa.2020.124376
|
| [50] |
A. L. Barabási, R. Albert, Emergence of scaling in random networks, Science, 286 (1999), 509. https://doi.org/10.1126/science.286.5439.509 doi: 10.1126/science.286.5439.509
|
| [51] |
N. Nakagiri, Y. Sakisaka, T. Togashi, S. Morita, K. Tainaka, Effects of habitat destruction in model ecosystems: parity law depending on species richness, Ecol. Inf., 5 (2010), 241–247. https://doi.org/10.1016/j.ecoinf.2010.05.003 doi: 10.1016/j.ecoinf.2010.05.003
|

Figures(6) / Tables(3)

Nariyuki Nakagiri, Hiroki Yokoi, Yukio Sakisaka, Kei-ichi Tainaka, Kazunori Sato. Influence of network structure on infectious disease control[J]. Mathematical Biosciences and Engineering, 2025, 22(4): 943-961. doi: 10.3934/mbe.2025034







 DownLoad:
DownLoad: