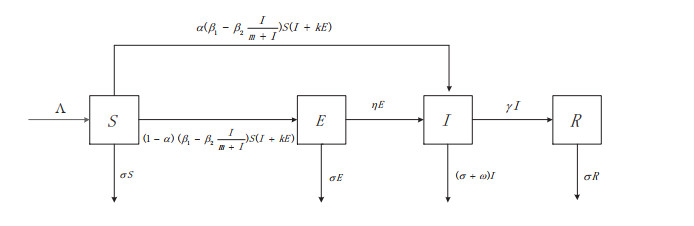
Media coverage can greatly impact the spread of infectious diseases. Taking into consideration the impacts of media coverage, we propose an SEIR model with a media coverage mediated nonlinear infection force. For this novel disease model, we identify the basic reproduction number using the next generation matrix method and establish the global threshold results: If the basic reproduction number $ \mathcal{R}_{0} < 1 $, then the disease-free equilibrium $ P_{0} $ is stable, and the disease dies out. If $ \mathcal{R}_{0} > 1 $, then the endemic equilibrium $ P^{*} $ is stable, and the disease persists. Sensitivity analysis indicates that the basic reproduction number $ \mathcal{R}_{0} $ is most sensitive to the population recruitment rate $ \Lambda $ and the disease transmission rate $ \beta _{1} $.
Citation: Jingli Xie, Hongli Guo, Meiyang Zhang. Dynamics of an SEIR model with media coverage mediated nonlinear infectious force[J]. Mathematical Biosciences and Engineering, 2023, 20(8): 14616-14633. doi: 10.3934/mbe.2023654
Media coverage can greatly impact the spread of infectious diseases. Taking into consideration the impacts of media coverage, we propose an SEIR model with a media coverage mediated nonlinear infection force. For this novel disease model, we identify the basic reproduction number using the next generation matrix method and establish the global threshold results: If the basic reproduction number $ \mathcal{R}_{0} < 1 $, then the disease-free equilibrium $ P_{0} $ is stable, and the disease dies out. If $ \mathcal{R}_{0} > 1 $, then the endemic equilibrium $ P^{*} $ is stable, and the disease persists. Sensitivity analysis indicates that the basic reproduction number $ \mathcal{R}_{0} $ is most sensitive to the population recruitment rate $ \Lambda $ and the disease transmission rate $ \beta _{1} $.

| [1] |
J. M. Tchuenche, N. Dube, C. P. Bhunu, R. J. Smith, C. T. Bauch, The impact of media coverage on the transmission dynamics of human influenza, BMC Public Health, 11 (2011), 1–14. https://doi.org/10.1186/1471-2458-11-S1-S5 doi: 10.1186/1471-2458-11-S1-S5

|
| [2] |
W. K. Zhou, Y. N. Xiao, J. M. Heffernan, Optimal media reporting intensity on mitigating spread of an emerging infectious disease, PLoS One, 14 (2019), e0213898. https://doi.org/10.1371/journal.pone.0213898 doi: 10.1371/journal.pone.0213898
|
| [3] |
R. S. Liu, J. H. Wu, H. P. Zhu, Media/psychological impact on multiple outbreaks of emerging infectious diseases, Comput. Math. Methods Med., 8 (2007), 153–164. https://doi.org/10.1080/17486700701425870 doi: 10.1080/17486700701425870
|
| [4] |
D. M. Xiao, S. G. Ruan, Global analysis of an epidemic model with nonmonotone incidence rate, Math. Biosci., 208 (2007), 419–429. https://doi.org/10.1016/j.mbs.2006.09.025 doi: 10.1016/j.mbs.2006.09.025
|
| [5] |
J. A. Cui, X. Tao, H. P. Zhu, An SIS infection model incorporating media coverage, Rocky Mt. J. Math., 38 (2008), 1323–1334. https://doi.org/10.1216/RMJ-2008-38-5-1323 doi: 10.1216/RMJ-2008-38-5-1323
|
| [6] | J. Cui, Z. Wu, An SIRS model for assessing impact of media coverage, Abstr. Appl. Anal., 2014 (2014). https://doi.org/10.1155/2014/424610 |
| [7] |
Y. Wang, J. Cao, Z. Jin, H. Zhang, G. Q. Sun, Impact of media coverage on epidemic spreading in complex networks, Physica A, 392 (2013), 5824–5835. https://doi.org/10.1016/j.physa.2013.07.067 doi: 10.1016/j.physa.2013.07.067
|
| [8] |
S. R. Gani, S. V. Halawar, Optimal control for the spread of infectious disease: the role of awareness programs by media and antiviral treatment, Optim. Control Appl. Methods, 39 (2018), 1407–1430. https://doi.org/10.1002/oca.2418 doi: 10.1002/oca.2418
|
| [9] |
A. Kumar, P. K. Srivastava, Y. P. Dong, Y. Takeuchi, Optimal control of infectious disease: information-induced vaccination and limited treatment, Physica A, 542 (2020), 123196. https://doi.org/10.1016/j.physa.2019.123196 doi: 10.1016/j.physa.2019.123196
|
| [10] |
S. S. Shanta, M. H. A. Biswas, The impact of media awareness in controlling the spread of infectious diseases in terms of SIR model, Math. Modell. Eng. Probl., 7 (2020), 368–376. https://doi.org/10.18280/mmep.070306 doi: 10.18280/mmep.070306
|
| [11] |
W. Wang, X. Q. Zhao, Threshold dynamics for compartmental epidemic models in periodic environments, J. Dyn. Differ. Equations, 20 (2008), 699–717. https://doi.org/10.1007/s10884-008-9111-8 doi: 10.1007/s10884-008-9111-8
|
| [12] | W. Xing, J. F. Gao, Q. S. Yan, Q. H. Zhou, Z. H. Yang, An epidemic model with saturated media/psychological impact (in Chinese), J. Northwest Univ. (Nat. Sci. Ed.), 48 (2018), 639–643. Available from: https://kns.cnki.net/kcms/detail/61.1072.N.20181015.1716.006.html. |
| [13] |
Q. L. Yan, S. Y. Tang, S. Gabriele, J. H. Wu, Media coverage and hospital notifications: correlation analysis and optimal media impact duration to manage a pandemic, J. Theor. Biol., 390 (2016), 1–13. https://doi.org/10.1016/j.jtbi.2015.11.002 doi: 10.1016/j.jtbi.2015.11.002
|
| [14] | Q. L. Yan, Y. L. Tang, D. D. Yan, J. Y. Wang, L. Q. Yang, X. P. Yang, et al., Impact of media reports on the early spread of COVID-19 epidemic, J. Theor. Biol., 502 (2020). https://doi.org/10.1016/j.jtbi.2020.110385 |
| [15] |
I. Ghosh, P. K. Tiwari, S. Samanta, I. M. Elmojtaba, N. Al-Salti, J. Chattopadhyay, A simple SI-type model for HIV/AIDS with media and self-imposed psychological fear, J. Math. Biosci., 306 (2018), 160–169. https://doi.org/10.1016/j.mbs.2018.09.014 doi: 10.1016/j.mbs.2018.09.014
|
| [16] |
S. Latifah, D. Aldila, W. Giyarti, H. Tasman, Mathematical study for an infectious disease with awareness-based SIS-M model, J. Phys. Conf. Ser., 1747 (2021), 012017. https://doi.org/10.1088/1742-6596/1747/1/012017 doi: 10.1088/1742-6596/1747/1/012017
|
| [17] |
G. O. Agaba, Y. N. Kyrychko, K. B. Blyuss, Mathematical model for the impact of awareness on the dynamics of infectious diseases, Math. Biosci., 286 (2017), 22–30. https://doi.org/10.1016/j.mbs.2017.01.009 doi: 10.1016/j.mbs.2017.01.009
|
| [18] |
P. van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. https://doi.org/10.1016/S0025-5564(02)00108-6 doi: 10.1016/S0025-5564(02)00108-6
|
| [19] | C. Castillo-Chaves, H. R. Thieme, Asymptotically autonomous epidemic models, in Mathematical Population Dynamics: Analysis of Heterogeneity. Volume One, Theory of Epidemics, (1995), 33–50. Available from: https://www.researchgate.net/publication/221711274. |
| [20] |
M. Y. Li, H. Smith, L. C. Wang, Global dynamics of an SEIR epidemic model with vertical transmission, SIAM J. Appl. Math., 62 (2001), 58–69. https://doi.org/10.1137/S0036139999359860 doi: 10.1137/S0036139999359860
|
| [21] |
N. Chitnis, J. M. Hyman, J. M. Cushing, Determining important parameters in the spread of malaria through the sensitivity analysis of a mathematical model, Bull. Math. Biol., 70 (2008), 1272–1296. https://doi.org/10.1007/s11538-008-9299-0 doi: 10.1007/s11538-008-9299-0
|
| [22] | Y. N. Sun, Analysis of Several Models on the Infectious Diseases with the Effect of Awareness (in Chinese), Master's thesis, North University of China, 2021. https://doi.org/10.27470/d.cnki.ghbgc.2021.001026 |
| [23] |
B. Tang, F. Xia, S. Y. Tang, N. L. Bragazzi, Q. Li, X. D. Sun, et al., The effectiveness of quarantine and isolation determine the trend of the COVID-19 epidemics in the final phase of the current outbreak in China, Int. J. Infect. Dis., 95 (2020), 288–293. https://doi.org/10.1016/j.ijid.2020.03.018 doi: 10.1016/j.ijid.2020.03.018
|
| [24] |
M. Mandal, S. Jana, S. K. Nandi, A. Khatua, S. Adak, T. K. Kar, A model based study on the dynamics of COVID-19: prediction and control, Chaos, Solitons Fractals, 136 (2020), 109889. https://doi.org/10.1016/j.chaos.2020.109889 doi: 10.1016/j.chaos.2020.109889
|
| [25] |
B. Tang, X. Wang, Q. Li, N. L. Bragazzi, S. Y. Tang, Y. N. Xiao, et al., Estimation of the transmission risk of the 2019-nCoV and its implication for public health interventions, J. Clin. Med., 9 (2020), 462. https://doi.org/10.3390/jcm9020462 doi: 10.3390/jcm9020462
|







Figures(8) / Tables(1)

Jingli Xie, Hongli Guo, Meiyang Zhang. Dynamics of an SEIR model with media coverage mediated nonlinear infectious force[J]. Mathematical Biosciences and Engineering, 2023, 20(8): 14616-14633. doi: 10.3934/mbe.2023654







 DownLoad:
DownLoad: