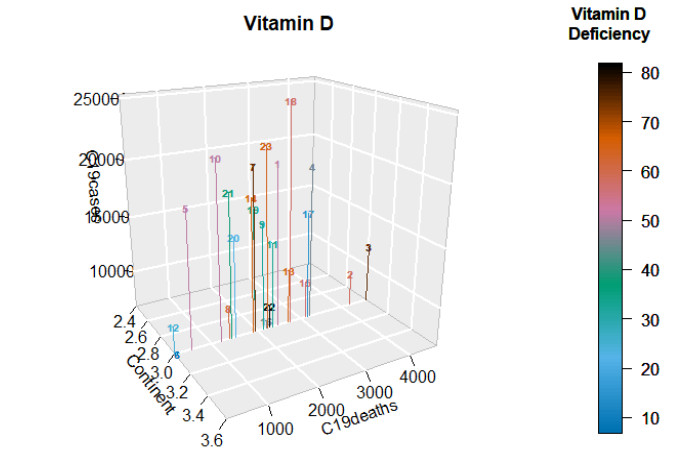
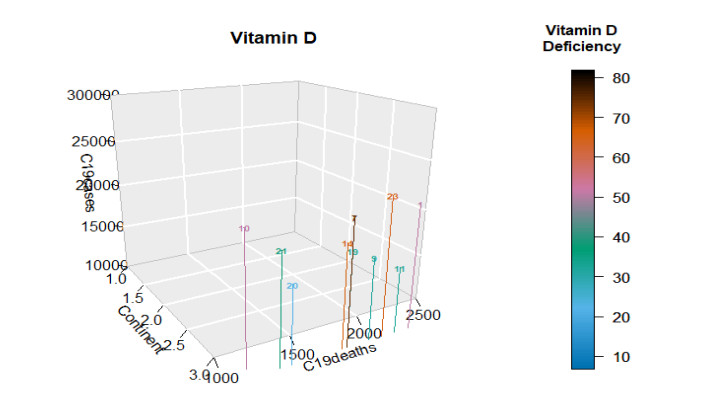
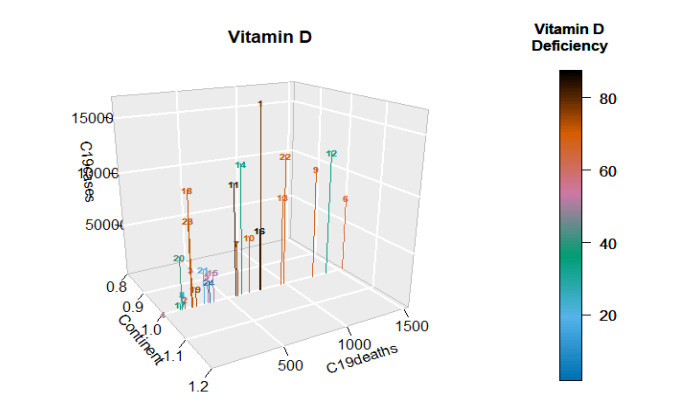
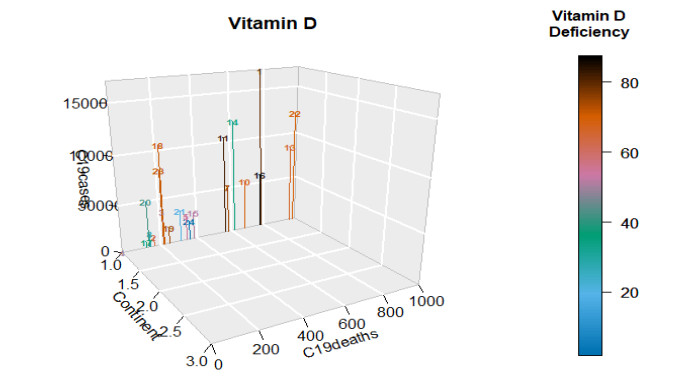
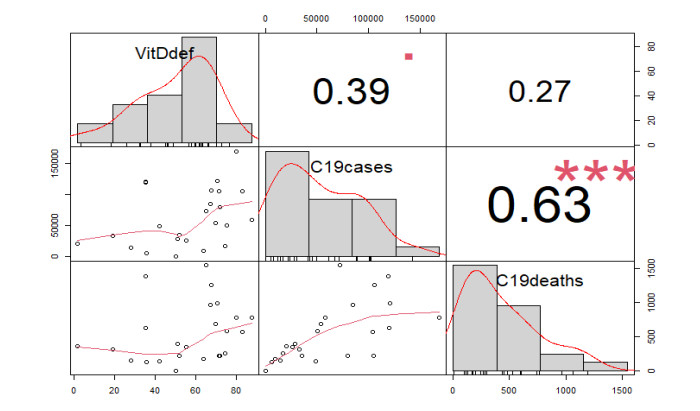
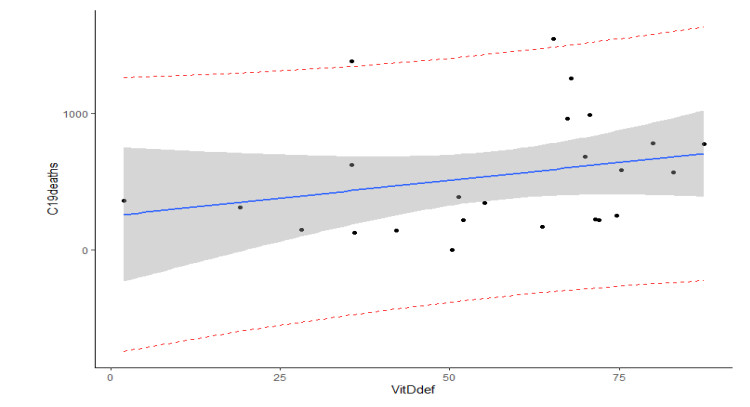
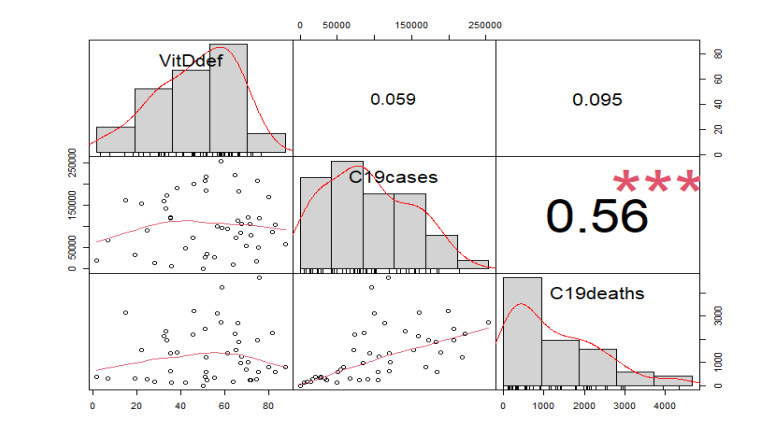
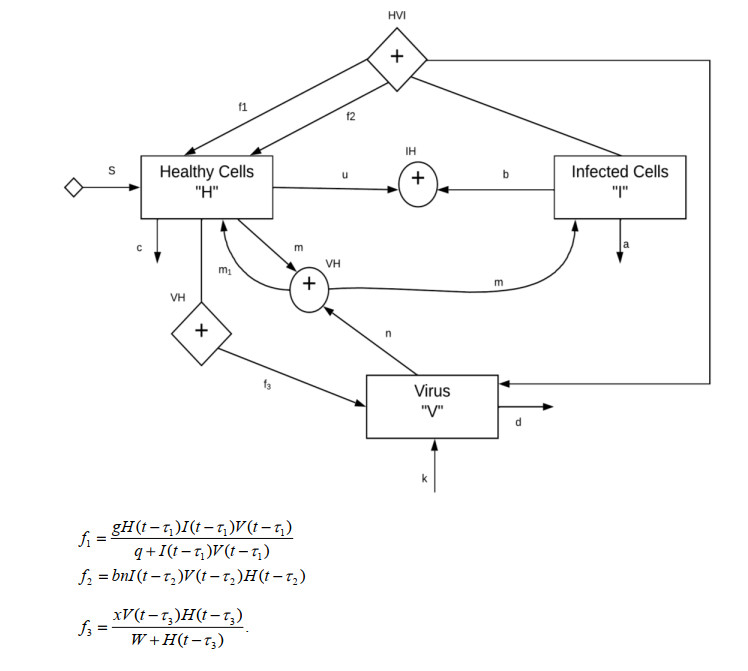
This paper presents some recent views on the aspects of vitamin D levels in relation to the COVID-19 infections and analyzes the relationship between the prevalence rates of vitamin D deficiency and COVID-19 death rates per million of various countries in Europe and Asia using the data from the PubMed database. The paper also discusses a new mathematical model of time-delay interactions between the body's immune healthy cells, infected cells, and virus particles with the effect of vitamin D levels. The model can be used to monitor the timely progression of healthy immune cells with the effects of the levels of vitamin D and probiotics supplement. It also can help to predict when the infected cells and virus particles free state can ever be reached as time progresses. The consideration of the time delay in the modeling due to effects of the infected cells or virus particles and the growth of healthy cells is also an important factor that can significantly change the outcomes of the body's immune cells as well as the infections.
Citation: Hoang Pham. Analyzing the relationship between the vitamin D deficiency and COVID-19 mortality rate and modeling the time-delay interactions between body's immune healthy cells, infected cells, and virus particles with the effect of vitamin D levels[J]. Mathematical Biosciences and Engineering, 2022, 19(9): 8975-9004. doi: 10.3934/mbe.2022417
This paper presents some recent views on the aspects of vitamin D levels in relation to the COVID-19 infections and analyzes the relationship between the prevalence rates of vitamin D deficiency and COVID-19 death rates per million of various countries in Europe and Asia using the data from the PubMed database. The paper also discusses a new mathematical model of time-delay interactions between the body's immune healthy cells, infected cells, and virus particles with the effect of vitamin D levels. The model can be used to monitor the timely progression of healthy immune cells with the effects of the levels of vitamin D and probiotics supplement. It also can help to predict when the infected cells and virus particles free state can ever be reached as time progresses. The consideration of the time delay in the modeling due to effects of the infected cells or virus particles and the growth of healthy cells is also an important factor that can significantly change the outcomes of the body's immune cells as well as the infections.

| [1] | Available online: https://patch.com/new-jersey/oceancity/nj-coronavirus-update-gov-murphy-considers-curfew-31-new-cases (accessed on16 March 2020). |
| [2] | Available online: https://www.osha.gov/SLTC/covid-19/medicalinformation.html (accessed on.17 March 2020). |
| [3] | Centers for Disease Control and Prevention, https://www.cdc.gov/coronavirus/2019-ncov/prevent-getting-sick/social-distancing.html (accessed on 8 April 2020) |
| [4] |
H. Pham, On estimating the number of deaths related to Covid-19, Mathematics, 8 (2020), 655. https://doi.org/10.3390/math8050655 doi: 10.3390/math8050655

|
| [5] | Worldometers, 2022 (accessed on April 1, 2022). |
| [6] | https://www.worldometers.info/coronavirus/?utm_campaign = homeAdvegas1?#countries |
| [7] | World Health Organization, https://covid19.who.int/ (accessed on April 1, 2022). |
| [8] |
H. Pham, Estimating the COVID-19 death toll by considering the time-dependent effects of various pandemic restrictions, Mathematics, 8 (2020), 1628. https://doi.org/10.3390/math8091628 doi: 10.3390/math8091628
|
| [9] |
A. W. H. Chin, J. T. S. Chu, M. R. A. Perera, K. P. Y. Hui, H. L. Yen, M. C. W. Chan, et al., Stability of SARS-CoV-2 in different environmental conditions, Lancet Microbe, 1 (2020), 1. https://doi.org/10.1016/S2666-5247(20)30003-3 doi: 10.1016/S2666-5247(20)30003-3
|
| [10] |
E. Dong, H. Du, L. Gardner, An interactive web-based dashboard to track COVID-19 in real time, Lancet Infect. Dis. 20 (2020), 533-534. https://doi.org/10.1016/S1473-3099(20)30120-1 doi: 10.1016/S1473-3099(20)30120-1
|
| [11] |
K. Prem, Y. Liu, T. W. Russell, A. J. Kucharski, R. M. Eggo, N. Davies, et al., The effect of control strategies to reduce social mixing on outcomes of the COVID-19 epidemic in Wuhan, China: A modelling study, Lancet Public Health, 20 (2020), e261-e270. https://doi.org/10.1016/S2468-2667(20)30073-6 doi: 10.1016/S2468-2667(20)30073-6
|
| [12] |
G. Sebastiani, M. Massa, E. Riboli, Covid-19 epidemic in Italy: Evolution, projections and impact of government measures, Eur. J. Epidemiol., 35 (2020), 341-345. https://doi.org/10.1007/s10654-020-00631-6 doi: 10.1007/s10654-020-00631-6
|
| [13] |
G. Onder, G. Rezza, S. Brusaferro, Case-Fatality rate and characteristics of patients dying in relation to COVID-19 in Italy, JAMA, 20 (2020), 1775-1776. https://doi.org/10.1001/jama.2020.4683 doi: 10.1001/jama.2020.4683
|
| [14] |
D. D. Rajgor, M. H. Lee, S. Archuleta, N. Bagdasarian, S. C. Quek, The many estimates of the COVID-19 case fatality rate, Lancet Infect. Dis., 20 (2020), 776-777. https://doi.org/10.1016/S1473-3099(20)30244-9 doi: 10.1016/S1473-3099(20)30244-9
|
| [15] |
A. J. Kucharski, T. W. Russell, C. Diamond, Y. Liu, J. Edmunds, S. Funk, et al., Early dynamics of transmission and control of COVID-19: A mathematical modelling study, Lancet Infect. Dis., 20 (2020), 553-558. https://doi.org/10.1016/S1473-3099(20)30144-4 doi: 10.1016/S1473-3099(20)30144-4
|
| [16] |
X. Wang, Y. Zhang, F. Fang, Role of vitamin D in COVID-19 infections and deaths, J. Evid. Based Med., 14 (2021), 5-6. https://doi.org/10.1111/jebm.12421 doi: 10.1111/jebm.12421
|
| [17] |
E. D. Gorham, C. F. Garland, F. C. Garland, W. B. Grant, S. B. Mohr, M. Lipkin, et al., Optimal vitamin D status for colorectal cancer prevention: A quantitative meta analysis, Am. J. Prev. Med., 32 (2007), 210-216. https://doi.org/10.1016/j.amepre.2006.11.004 doi: 10.1016/j.amepre.2006.11.004
|
| [18] |
A. G. Pittas, S. S. Harris, P. C. Stark, H. B. Dawson, The effects of calcium and vitamin D supplementation on blood glucose and markers of inflammation in nondiabetic adults, Diabetes Care, 30 (2007), 980-986. https://doi.org/10.2337/dc06-1994 doi: 10.2337/dc06-1994
|
| [19] |
M. Urashima, T. Segawa, M. Okazaki, M. Kurihara, Y. Wada, H. Ida, Randomized trial of vitamin D supplementation to prevent seasonal influenza A in schoolchildren, Am. J. Clin. Nutr., 91 (2010), 1255-1260. https://doi.org/10.3945/ajcn.2009.29094 doi: 10.3945/ajcn.2009.29094
|
| [20] |
H. Pham, D. H. Pham, A novel generalized logistic dependent model to predict the presence of breast cancer based on biomarkers, Concurr. Comput. Pract. Exper., 32 (2020), 1. https://doi.org/10.1002/cpe.5467 doi: 10.1002/cpe.5467
|
| [21] |
V. Verhoeven, K. Vanpuyenbroeck, M. Lopez-Hartmann, J. Wens, R. Remmen, Walk on the sunny side of life—epidemiology of hypovitaminosis D and mental health in elderly nursing home residents, J. Nutr. Health Aging, 16 (2012), 417-420. https://doi.org/10.1007/s12603-011-0361-5 doi: 10.1007/s12603-011-0361-5
|
| [22] |
D. Boersma, O. Demontiero, A. Z. Mohtasham, S. Hassan, H. Suarez, D. Geisinger, et al., Vitamin D status in relation to postural stability in the elderly, J. Nutr. Health Aging, 16 (2012), 270-275. https://doi.org/10.1007/s12603-011-0345-5 doi: 10.1007/s12603-011-0345-5
|
| [23] |
N. Charoenngam, M. F. Holick, Immunologic effects of vitamin D on human health and disease, Nutrients, 12 (2020), 2097. https://doi.org/10.3390/nu12072097 doi: 10.3390/nu12072097
|
| [24] |
A. R. Martineau, D. A. Jolliffe, L. Greenberg, J. F. Aloia, P. Bergman, G. Dubnov-Raz, Vitamin D supplementation to prevent acute respiratory infections: Individual participant data meta-analysis, Health Technol. Assess., 23 (2019), 23020. https://doi.org/10.3310/hta23020 doi: 10.3310/hta23020
|
| [25] | C. E. Hayes, J. M. Ntambi, Multiple sclerosis: Lipids, Lymphocytes, and vitamin D, Immunometabolism, 10 (2020), 19. |
| [26] |
K. de Haan, A. B. J. Groeneveld, H. R. H. de Geus, M. Egal, A. Struijs, Vitamin D deficiency as a risk factor for infection, sepsisand mortality in the critically ill: Systematic review and meta-analysis, Crit. Care, 66 (2014), 6604. https://doi.org/10.1186/s13054-014-0660-4 doi: 10.1186/s13054-014-0660-4
|
| [27] |
A. Braun, D. Chang, K. Mahadevappa, F. K. Gibbons, Y. Liu, E. Giovannucci, Association of low serum 25-hydroxyvitamin D levels and mortality in the critically ill, Crit. Care Med., 39 (2011), 671. https://doi.org/10.1097/CCM.0b013e318206ccdf doi: 10.1097/CCM.0b013e318206ccdf
|
| [28] |
P. Autier, M. Boniol, C. Pizot, P. Mullie, Vitamin D status and ill health: A systematic review, Lancet Diabetes Endocrinol., 2 (2014) 76. https://doi.org/10.1016/S2213-8587(13)70165-7 doi: 10.1016/S2213-8587(13)70165-7
|
| [29] |
Y. F. Zhou, B. A. Luo, L. L. Qin, The association between vitamin D deficiency and community-acquired pneumonia, Medicine, 98 (2019), e17252. https://doi.org/10.1097/MD.0000000000017252 doi: 10.1097/MD.0000000000017252
|
| [30] |
E. C. Goodall, A. C. Granados, K. Luinstra, E. Pullenayegum, B. L. Coleman, M. Loeb, Vitamin D3 and gargling for the prevention of upper respiratory tract infections: A randomized controlled trial, BMC Infect. Dis., 11 (2014), 14273. https://doi.org/10.1186/1471-2334-14-273 doi: 10.1186/1471-2334-14-273
|
| [31] |
A. S. Vanherwegen, C. Gysemans, C. Mathieu, Regulation of immune function by vitamin D and its use in diseases of immunity, Endocrinol. Metab. Clin. N. Am., 46 (2017), 1061. https://doi.org/10.1016/j.ecl.2017.07.010 doi: 10.1016/j.ecl.2017.07.010
|
| [32] |
C. Greiller, A. Martineau, Modulation of the immune response to respiratory viruses by vitamin D, Nutrients, 7 (2015), 4240-4270. https://doi.org/10.3390/nu7064240 doi: 10.3390/nu7064240
|
| [33] |
M. T. Zdrenghea, H. Makrinioti, C. Bagacean, A. Bush, S. L. Johnston, L.A. Stanciu, Vitamin D modulation of innate immune responses to respiratory viral infections, Rev. Med. Virol., 27 (2016), e1909. https://doi.org/10.1002/rmv.1909 doi: 10.1002/rmv.1909
|
| [34] |
J. R. Sabetta, P. DePetrillo, R. J. Cipriani, J. Smardin, L. A. Burns, M. L. Landry, Serum 25-hydroxyvitamin d and the incidence of acute viral respiratory tract infections in healthy adults, PLoS One, 5 (2010), e11088. https://doi.org/10.1371/journal.pone.0011088 doi: 10.1371/journal.pone.0011088
|
| [35] | T. R. Ingham, B. Jones, C. A. Camargo, J. Kirman, A. C. Dowell, J. Crane, Association of vitamin D deficiency with severity of acute respiratory infection: A case-control study in New Zealand children, Eur. Respir. J. Eur. Respir. Soc., 44 (2014), 124. |
| [36] |
C. FGunville, P. MMourani, A. AGinde, The role of vitamin D in prevention and treatment of infection, Inflamm. Allergy-DrugTargets, 12 (2013), 239-245. https://doi.org/10.2174/18715281113129990046 doi: 10.2174/18715281113129990046
|
| [37] |
A. L. Khoo, L. Chai, H. Koenen, I. Joosten, M. Netea, A. van der Ven, Translating the role of vitamin D3 in infectious diseases, Crit. Rev. Microbiol., 38 (2012), 122. https://doi.org/10.3109/1040841X.2011.622716 doi: 10.3109/1040841X.2011.622716
|
| [38] |
R. Taha, S. Abureesh, S. Alghamdi, R. Y. Hassan, M. M. Cheikh, R. A. Bagabir, The relationship between vitamin D and infections including COVID-19: Any hopes? Int. J. Gen. Med., 14 (2021), 3849. https://doi.org/10.2147/IJGM.S317421 doi: 10.2147/IJGM.S317421
|
| [39] |
A. R. Martineau, D. A. Jolliffe, R. L. Hooper, L. Greenberg, J. F. Aloia, P. Bergman, Vitamin D supplementation to prevent acute respiratory tract infections: Systematic review and meta-analysis of individual participant data, BMJ, 65 (2017), 6583. https://doi.org/10.1136/bmj.i6583 doi: 10.1136/bmj.i6583
|
| [40] |
R. C. A. Dancer, D. Parekh, S. Lax, V. D'Souza, S. Zheng, C. R. Bassford, Vitamin D deficiency contributes directly to the acute respiratory distress syndrome (ARDS), Thorax BMJ, 70 (2015), 617. https://doi.org/10.1136/thoraxjnl-2014-206680 doi: 10.1136/thoraxjnl-2014-206680
|
| [41] |
D. R. Thickett, T. Moromizato, A. A. Litonjua, K. Amrein, S. A. Quraishi, K. A. Lee-Sarwar, Association between prehospital vitamin D status and incident acute respiratory failure in critically ill patients: A retrospective cohort study, BMJ Open Respir. Res., 2 (2015), e74. https://doi.org/10.1136/bmjresp-2014-000074 doi: 10.1136/bmjresp-2014-000074
|
| [42] | M. T. Cantorna, B. D. Mahon, D-hormone and the immune system, J. Rheumatol. Suppl., 76 (2005), 11-20. |
| [43] |
A. Antico, M. Tampoia, R. Tozzoli, N. Bizzaro, Can supplementation with vitamin D reduce the risk or modify the course of autoimmune diseases? A systematic review of the literature, Autoimmun. Rev., 12 (2012), 127. https://doi.org/10.1016/j.autrev.2012.07.007 doi: 10.1016/j.autrev.2012.07.007
|
| [44] | T. Michigami, O. Rickets, Consensus on vitamin D deficiency and insufficiency in children, Clin. Calcium., 28 (2018), 1307-1311. |
| [45] |
W. Grant, H. Lahore, S. McDonnell, C. Baggerly, C. French, J. Aliano, Evidence that vitamin D supplementation could reduce risk of influenza and COVID-19 infections and deaths, Nutrients, 12 (2020), 988. https://doi.org/10.3390/nu12040988 doi: 10.3390/nu12040988
|
| [46] |
P. A. Danai, S. Sinha, M. Moss, M. J. Haber, G. S. Martin, Seasonal variation in the epidemiology of sepsis, Crit. Care Med., 35 (2007), 410-415. https://doi.org/10.1097/01.CCM.0000253405.17038.43 doi: 10.1097/01.CCM.0000253405.17038.43
|
| [47] |
W. B. Grant, Variations in vitamin D production could possibly explain the seasonality of childhood respiratory infections in Hawaii, Pediatr. Infect. Dis. J., 27 (2008), 853. https://doi.org/10.1097/INF.0b013e3181817bc1 doi: 10.1097/INF.0b013e3181817bc1
|
| [48] |
J. J. Cannell, R. Vieth, J. Umhau, M. Holick, W. Grant, S. Madronich, et al., Epidemic influenza and vitamin D, Epidemiol. Infect., 134 (2006), 1129-1140. https://doi.org/10.1017/S0950268806007175 doi: 10.1017/S0950268806007175
|
| [49] |
V. Dimitrov, C. Barbier, A. Ismailova, Y. Wang, K. Dmowski, R. Salehi-Tabar, et al., Vitamin D-regulated gene expression profiles: species-specificity and cell-specific effects on metabolism and immunity, Endocrinology, 162 (2021), 218. https://doi.org/10.1210/endocr/bqaa218 doi: 10.1210/endocr/bqaa218
|
| [50] |
M. Olliver, L. Spelmink, J. Hiew, U. Meyer-Hoffert, B. Henriques-Normark, P. Bergman, Immunomodulatory effects of vitamin D on innate and adaptive immune responses to Streptococcus pneumoniae, J. Infect. Dis., 208 (2013), 1474-1481. https://doi.org/10.1093/infdis/jit355 doi: 10.1093/infdis/jit355
|
| [51] |
S. Maggini, E. S. Wintergerst, S. Beveridge, D. H. Hornig, Selected vitamins and trace elements support immune function by strengthening epithelial barriers and cellular and humoral immune responses, Br. J. Nut., 98 (2007), S29-S35. https://doi.org/10.1017/S0007114507832971 doi: 10.1017/S0007114507832971
|
| [52] |
W. B. Grant, H. Lahore, S. L. McDonnell, C. A. Baggerly, C. B. French, J. L. Aliano, et al., Evidence that vitamin D supplementation could reduce risk of influenza and COVID-19 infections and deaths, Nutrients, 12 (2020), 988. https://doi.org/10.3390/nu12040988 doi: 10.3390/nu12040988
|
| [53] |
S. Piumika, T. J. Dhanushya, K. Neil, J. Ranil, Impact of vitamin D deficiency on COVID-19, Clin. Nutr. ESPEN, 44 (2021), 372-378. https://doi.org/10.1016/j.clnesp.2021.05.011 doi: 10.1016/j.clnesp.2021.05.011
|
| [54] | E. Laird, R. A. Kenny, Vitamin D deficiency in Ireland-implications for COVID-19. Results from the Irish Longitudinal Study on Ageing (TILDA), April 2020. https://doi.org/10.38018/TildaRe.2020-05 |
| [55] |
D. P. Richardson, J. A. Lovegrove, Nutritional status of micronutrients as a possible and modifiable risk factor for COVID-19: A UK perspective, Br. J. Nutr., 125 (2021), 678-684. https://doi.org/10.1017/S000711452000330X doi: 10.1017/S000711452000330X
|
| [56] |
M. Ebadi, A. J. Montano-Loza, Perspective: Improving vitamin D status in the management of COVID-19, Eur. J. Clin. Nutr., 74 (2020), 856-859. https://doi.org/10.1038/s41430-020-0661-0 doi: 10.1038/s41430-020-0661-0
|
| [57] |
W. B. Grant, H. Lahore, S. L. McDonnell, C. A. Baggerly, C. B. French, J. L. Aliano, et al., Evidence that vitamin D supplementation could reduce risk of influenza and COVID-19 infections and deaths, Nutrients, 12 (2020), 988. https://doi.org/10.3390/nu12040988 doi: 10.3390/nu12040988
|
| [58] | D. M. McCartney, D. G. Byrne, Optimisation of vitamin D status for enhanced Immuno-protection against Covid-19, Ir. Med. J., 113 (2020), 58. |
| [59] |
M. Di Rosa, M. Malaguarnera, F. Nicoletti, L. Malaguarnera, Vitamin D3: A helpful immuno-modulator. Immunology, 134 (2020), 123-139. https://doi.org/10.1111/j.1365-2567.2011.03482.x doi: 10.1111/j.1365-2567.2011.03482.x
|
| [60] |
J. Zhong, J. Tang, C. Ye, L. Dong, The immunology of COVID-19: Is immune modulation an option for treatment? Lancet Rheumatol., 2 (2020), e428-e436. https://doi.org/10.1016/S2665-9913(20)30120-X doi: 10.1016/S2665-9913(20)30120-X
|
| [61] |
A. Panarese, E. Shahini, Covid-19, and vitamin D, Aliment. Pharmacol. Ther., 51 (2020), 993. https://doi.org/10.1111/apt.15752 doi: 10.1111/apt.15752
|
| [62] |
G. Grasselli, T. Tonetti, A. Protti, T. Langer, M. Girardis, G. Bellani, et al., Pathophysiology of COVID-19-associated acute respiratory distress syndrome: A multicentre prospective observational study, Lancet Respir. Med., 8 (2020), 1201-1208. https://doi.org/10.1016/S2213-2600(20)30370-2 doi: 10.1016/S2213-2600(20)30370-2
|
| [63] |
A. A. Ginde, J. M. Mansbach, C. A. Camargo, Association between serum 25-hydroxyvitamin D level and upper respiratory tract infection in the Third National Health and Nutrition Examination Survey, Arch. Int. Med., 169 (2009), 384-390. https://doi.org/10.1001/archinternmed.2008.560 doi: 10.1001/archinternmed.2008.560
|
| [64] |
D. A. Jolliffe, C. J. Griffiths, A. R. Martineau, Vitamin D in the prevention of acute respiratory infection: Systematic review of clinical studies, J. Steroid Biochem. Mol. Biol., 136 (2020), 321-329. https://doi.org/10.1016/j.jsbmb.2012.11.017 doi: 10.1016/j.jsbmb.2012.11.017
|
| [65] |
D. Prantas, S. Nandeeta, O. A. Bright, E. H. John, P. Prince, M. Aliu, et al., Effect of vitamin D deficiency on COVID-19 status: A systematic review, COVID, 1 (2021), 97-104. https://doi.org/10.3390/covid1010008 doi: 10.3390/covid1010008
|
| [66] |
A. Radujkovic, T. Hippchen, S. Tiwari-Heckler, S. Dreher, M. Boxberger, U. Merle, Vitamin D deficiency and outcome of COVID-19 patients, Nutrients, 12 (2020), 2757. https://doi.org/10.3390/nu12092757 doi: 10.3390/nu12092757
|
| [67] |
D. O. Meltzer, T. J. Best, H. Zhang, T. Vokes, V. Arora, J. Solway, Association of vitamin D status and other clinical characteristics with COVID-19 test results, JAMA Netw. Open, 3 (2020), e2019722. https://doi.org/10.1001/jamanetworkopen.2020.19722 doi: 10.1001/jamanetworkopen.2020.19722
|
| [68] |
C. E. Hastie, D. F. Mackay, F. Ho, C. A. Celis-Morales, S. V. Katikireddi, C. L. Niedzwiedz, et al., Vitamin D concentrations and COVID-19 infection in UK Biobank, Diabetes Metab. Syndr. Clin. Res. Rev., 14 (2020), 561-565. https://doi.org/10.1016/j.dsx.2020.04.050 doi: 10.1016/j.dsx.2020.04.050
|
| [69] |
A. Abrishami, N. Dalili, P. M. Torbati, R. Asgari, M. Arab-Ahmadi, B. Behnam, et al., Possible association of vitamin D status with lung involvement and outcome in patients with COVID-19: A retrospective study, Eur. J. Nutr., 60 (2020), 2249-2257. https://doi.org/10.1007/s00394-020-02411-0 doi: 10.1007/s00394-020-02411-0
|
| [70] |
V. Baktash, T. Hosack, N. Patel, S. Shah, P. Kandiah, K. Van den Abbeele, et al., Vitamin D status and outcomes for hospitalised older patients with COVID-19, Postgrad. Med. J., 2 (2020), 1-6. https://doi.org/10.1136/postgradmedj-2020-138712 doi: 10.1136/postgradmedj-2020-138712
|
| [71] |
Z. Maghbooli, M. A. Sahraian, M. Ebrahimi, M. Pazoki, M. Kafan, S. Tabriz, et al., Vitamin D sufficiency, a serum 25-hydroxyvitamin D at least 30 ng/mL reduced risk for adverse clinical outcomes in patients with COVID-19 infection, PLoS One, 15 (2020), e0239799. https://doi.org/10.1371/journal.pone.0239799 doi: 10.1371/journal.pone.0239799
|
| [72] |
K. Ye, F. Tang, X. Liao, B. A. Shaw, M. Deng, G. Huang, et al., Does serum vitamin D level affect COVID-19 infection and its severity? A case-control study, J. Am. Coll. Nutr., (2020), 1-8. https://doi.org/10.1080/07315724.2020.1826005 doi: 10.1080/07315724.2020.1826005
|
| [73] |
P. Bergman, A. C. Norlin, S. Hansen, R. S. Rekha, B. Agerberth, L. Björkhem-Bergman, et al., Vitamin D3 supplementation in patients with frequent respiratory tract infections: A randomised and double-blind intervention study, BMJ Open, 2 (2012), e001663. https://doi.org/10.1136/bmjopen-2012-001663 doi: 10.1136/bmjopen-2012-001663
|
| [74] |
A. R. Martineau, N. G. Forouhi, Vitamin D for COVID-19: A case to answer? Lancet Diabetes Endocrinol, 8 (2020), 735-736. https://doi.org/10.1016/S2213-8587(20)30268-0 doi: 10.1016/S2213-8587(20)30268-0
|
| [75] |
N. Ali, Role of vitamin D in preventing of COVID-19 infection, progression and severity, J. Infect. Public Health, 13 (2020), 1373-1380. https://doi.org/10.1016/j.jiph.2020.06.021 doi: 10.1016/j.jiph.2020.06.021
|
| [76] |
P. C. Ilie, S. Stefanescu, L. Smith, The role of vitamin D in the prevention of coronavirus disease 2019 infection and mortality, Aging Clin. Exp. Res., 32 (2020), 1195-1198. https://doi.org/10.1007/s40520-020-01570-8 doi: 10.1007/s40520-020-01570-8
|
| [77] |
S. Walrand, Autumn COVID-19 surge dates in Europe correlated to latitudes, not to temperature-humidity, pointing to vitamin D as contributing factor, Sci. Rep., 11 (2021), 1981. https://doi.org/10.1038/s41598-021-81419-w doi: 10.1038/s41598-021-81419-w
|
| [78] |
E. Klingberg, G. Oleröd, J. Konar, M. Petzold, O. Hammarsten, Seasonal variations in serum 25-hydroxy vitamin D levels in a Swedish cohort, Endocrine, 49 (2015), 800-808. https://doi.org/10.1007/s12020-015-0548-3 doi: 10.1007/s12020-015-0548-3
|
| [79] | Institute of Medicine. Dietary reference intakes for calcium and vitamin D. Washington, DC: The National Academies Press, 2011. |
| [80] |
M. F. Holick N. C. Binkley, H. A. Bischoff-Ferrari, C. M. Gordon, D. A. Hanley, R. P. Heaney, et al., Evaluation, treatment, and prevention of vitamin D deficiency: An Endocrine Society Clinical Practice Guideline, J. Clin. Endocrinol. Metab., 96 (2011), 1911-1130. https://doi.org/10.1210/jc.2011-0385 doi: 10.1210/jc.2011-0385
|
| [81] |
D. E. Roth, P. Martz, R. Yeo, C. Prosser, M. Bell, A. B. Jones, Are national vitamin D guidelines sufficient to maintain adequate blood levels in children? Can. J. Public Health, 96 (2005), 443-449. https://doi.org/10.1007/BF03405185 doi: 10.1007/BF03405185
|
| [82] | D. Rucker, J. A. Allan, G. H. Fick, D. A. Hanley, Vitamin D insufficiency in a population of healthy western Canadians, CMAJ, 166 (2002), 1517-1524. |
| [83] |
R. Vieth, D. E. Cole, G. A. Hawker, H. M. Trang, L. A. Rubin, Winter time vitamin D insufficiency is common in young Canadian women, and their vitamin D intake does not prevent it, Eur. J. Clin. Nutr., 55 (2001), 1091-1097. https://doi.org/10.1038/sj.ejcn. 1601275 doi: 10.1038/sj.ejcn.1601275
|
| [84] |
R. P. Heaney, K. M. Davies, T. C. Chen, M. F. Holick, M. J. Barger-Lux, Human serum 25-hydroxycholecalciferol response to extended oral dosing with cholecalciferol, Am. J. Clin. Nutr., 77 (2003), 204-210. https://doi.org/10.1093/ajcn/77.1.204 doi: 10.1093/ajcn/77.1.204
|
| [85] |
J. N. Hathcock, A. Shao, R. Vieth, R. Heaney, Risk assessment for vitamin D, Am. J. Clin. Nutr., 85 (2007), 6-18. https://doi.org/10. 1093/ajcn/85.1.6 doi: 10.1093/ajcn/85.1.6
|
| [86] |
M. Pfeifer, B. Begerow, H. W. Minne, C. Abrams, D. Nachtigall, C. Hansen, Effects of a short-term vitamin D and calcium supplementation on body sway and secondary hyperparathyroidism in elderly women, J. Bone Miner Res., 15 (2000), 1113-1118. https://doi.org/10.1359/jbmr.2000.15.6.1113 doi: 10.1359/jbmr.2000.15.6.1113
|
| [87] | Medicine Io, Dietary reference intakes for calcium, phosphorus, magnesium, vitamin D, and fluoride. The Washington, DC: National Academies Press, 1997. |
| [88] |
J. F. Aloia, M. Patel, R. DiMaano, M. Li-Ng, S. A. Talwar, M. Mikhail, et al., Vitamin D intake to attain a desired serum 25- hydroxyvitamin D concentration, Am. J. Clin. Nutr., 87 (2008), 1952-1958. https://doi.org/10.1093/ajcn/87.6.1952 doi: 10.1093/ajcn/87.6.1952
|
| [89] |
A. C. Ross, J. E. Manson, S. A. Abrams, J. F. Aloia, P. M. Brannon, S. K. Clinton, et al., The 2011 report on dietary reference intakes for calcium and vitamin D from the institute of medicine: what clinicians need to know, J. Clin. Endocrinol. Metab., 96 (2011), 53-58. https://doi.org/10.1016/j.jada.2011.01.004 doi: 10.1016/j.jada.2011.01.004
|
| [90] | EFSA Panel on Dietetic Products N, Allergies. Scientific opinion on the tolerable upper intake level of vitamin D, EFSA J., 10 (2012), 2813. https://doi.org/10.2903/j.efsa.2012.2813 |
| [91] |
D. A. Hanley, A. Cranney, G. Jones, S. J. Whiting, W. D. Leslie, D. E. Cole, et al., Vitamin D in adult health and disease: A review and guideline statement from Osteoporosis Canada, CMAJ, 182 (2010), E610-E618. https://doi.org/10.1503/cmaj.080663 doi: 10.1503/cmaj.080663
|
| [92] |
S. M. Pietras, B. K. Obayan, M. H. Cai, M. F. Holick, Vitamin D2 treatment for vitamin D deficiency and insufficiency for up to 6 Years, JAMA Intern. Med., 169 (2009), 1806-1818. https://doi.org/10. 1001/archinternmed.2009.361 doi: 10.1001/archinternmed.2009.361
|
| [93] |
S. L. McDonnell, C. A. Baggerly, C. B. French, L. L. Baggerly, C. F. Garland, E. D. Gorham, et al., Breast cancer risk markedly lower with serum 25-hydroxyvitamin D concentrations ≥60 vs < 20 ng/ml (150 vs 50 nmol/L): Pooled analysis of two randomized trials and a prospective cohort, PLoS One, 13 (2018), e0199265. https://doi.org/10.1371/journal.pone.0199265 doi: 10.1371/journal.pone.0199265
|
| [94] |
J. M. Madden, L. Murphy, L. Zgaga, K. K. Bennett, De novo vitamin D supplement use post-diagnosis is associated with breast cancer survival, Breast Cancer Res. Treat., 172 (2018), 179-90. https://doi.org/10.1007/s10549-018-4896-6 doi: 10.1007/s10549-018-4896-6
|
| [95] |
N. Mirhosseini, H. Vatanparast, S. M. Kimball, The association between Serum 25(OH)D status and blood pressure in participants of a community-based program taking vitamin D supplements, Nutrients, 9 (2017), 1244. https://doi.org/10.3390/nu9111244 doi: 10.3390/nu9111244
|
| [96] |
A. Rusińska, P. Płudowski, M. Walczak, M. K. Borszewska-Kornacka, A. Bossowski, D. Chlebna-Sokół, et al., Vitamin D supplementation guidelines for general population and groups at risk of vitamin D deficiency in poland—recommendations of the polish society of pediatric endocrinology and diabetes and the expert panel with participation of national specialist consultants and representatives of scientific societies, Front. Endocrinol., 9 (2018). https://doi.org/10.3389/fendo.2018.00246 doi: 10.3389/fendo.2018.00246
|
| [97] |
K. Amrein, M. Scherkl, M. Hoffmann, S. Neuwersch-Sommeregger, M. Köstenberger, A. Tmava Berisha, et al., Vitamin D deficiency 2.0: An update on the current status worldwide, European J. Clin. Nutr., 74 (2020), 1498-1513. https://doi.org/10.1038/s41430-020-0558-y doi: 10.1038/s41430-020-0558-y
|
| [98] |
N. Charoenngam, M. F. Holick, Immunologic effects of vitamin d on human health and disease, Nutrients, 12 (2020), 1-28. https://doi.org/10.3390/nu12072097 doi: 10.3390/nu12072097
|
| [99] |
M. Chakhtoura, M. Rahme, N. Chamoun, G. El-Hajj Fuleihan, Vitamin D in the Middle East and North Africa, Bone Rep., 8 (2018), 135-146. https://doi.org/10.1016/j.bonr.2018.03.004 doi: 10.1016/j.bonr.2018.03.004
|
| [100] |
A. Batieha, Y. Khader, H. Jaddou, D. Hyassat, Z. Batieha, M. Khateeb, et al., Vitamin D status in Jordan: dress style and gender discrepancies, Ann. Nutr. Metab., 58 (2011), 10-18. https://doi.org/10.1159/000323097 doi: 10.1159/000323097
|
| [101] |
P. Bergman, A. U. Lindh, L. Björkhem-Bergman, J. D. Lindh, Vitamin D and respiratory tract infections: A systematic review and meta-analysis of randomized controlled trials, PLoS One, 8 (2013), e65835. https://doi.org/10.1371/journal.pone.0065835 doi: 10.1371/journal.pone.0065835
|
| [102] |
D. A. Jolliffe, C. A. Camargo, J. D. Sluyter, M. Aglipay, J. F. Aloia, D. Ganmaa, et al., Vitamin D supplementation to prevent acute respiratory infections: A systematic review and meta-analysis of aggregate data from randomised controlled trials, Lancet Diab. Endocrinol., 9 (2021), 276-292. https://doi.org/10.1136/thorax-2020-BTSabstracts.105 doi: 10.1136/thorax-2020-BTSabstracts.105
|
| [103] |
J. M. Rhodes, S. Subramanian, E. Laird, G. Griffig, R. A. Kenny, Perspective: Vitamin D deficiency and COVID-19 severity—plausibly linked by latitude, ethnicity, impacts on cytokines, ACE2 and thrombosis, J. Intern. Med., 289 (2021), 97-115. https://doi.org/10.1111/joim.13149 doi: 10.1111/joim.13149
|
| [104] |
D. O. Meltzer, T. J. Best, H. Zhang, T. Vokes, V. Arora, J. Solway, Association of vitamin D status and other clinical characteristics with COVID-19 test results, JAMA Netw. Open., 3 (2020), e2019722. https://doi.org/10.1001/jamanetworkopen.2020.19722 doi: 10.1001/jamanetworkopen.2020.19722
|
| [105] |
E. Merzon, D. Tworowski, A. Gorohovski, S. Vinker, A. G. Cohen, I. Green, et al., Low plasma 25(OH) vitamin D level is associated with increased risk of COVID-19 infection: An Israeli population-based study, FEBS J., 287 (2020), 3693-3702. https://doi.org/10.1111/febs.15495 doi: 10.1111/febs.15495
|
| [106] |
A. Jain, R. Chaurasia, N. S. Sengar, M. Singh, S. Mahor, S, Narain, Analysis of vitamin D level among asymptomatic and critically ill COVID-19 patients and its correlation with inflammatory markers, Sci. Rep., 10 (2020), 20191. https://doi.org/10.1038/s41598-020-77093-z doi: 10.1038/s41598-020-77093-z
|
| [107] | D. De Smet, K. De Smet, P. Herroelen, S. Gryspeerdt, G. A. Martens, Vitamin D deficiency as risk factor for severe COVID-19: A convergence of two pandemics, medRxiv, (2020). https://doi.org/10.1101/2020.05.01.20079376 |
| [108] |
R. L. Schleicher, M. R. Sternberg, A. C. Looker, E. A. Yetley, D. A. Lacher, C. T. Sempos, et al., National estimates of serum total 25-Hydroxyvitamin D and metabolite concentrations measured by liquid chromatography-Tandem mass spectrometry in the US population during 2007-2010, J. Nutr., 146 (2016), 1051-1061.https://doi.org/10.3945/jn.115.227728 doi: 10.3945/jn.115.227728
|
| [109] |
K. Sarafin, R. Durazo-Arvizu, L. Tian, K. W. Phinney, S. Tai, J. E. Camara, et al., Standardizing 25-hydroxyvitamin D values from the Canadian health measures survey, Am. J. Clin. Nutr., 102 (2015), 1044-1050. https://doi.org/10.3945/ajcn.114.103689 doi: 10.3945/ajcn.114.103689
|
| [110] |
K. D. Cashman, K. G. Dowling, Z. Škrabáková, M. Gonzalez-Gross, J. Valtueña, S. De Henauw, et al., Vitamin D deficiency in Europe: pandemic? Am. J. Clin. Nutr., 103 (2016), 1033-1044. https://doi.org/ 10.3945/ajcn.115.120873 doi: 10.3945/ajcn.115.120873
|
| [111] |
K. D. Cashman, Vitamin D deficiency: Defining, prevalence, causes, and strategies of addressing, Calcif. Tissue Int., (2019). https://doi.org/10.1007/s00223-019-00559-4 doi: 10.1007/s00223-019-00559-4
|
| [112] |
H. Pham, Modeling U.S. mortality and risk-cost optimization on life expectancy, IEEE Trans. Reliabil., 60 (2011), 125-133. https://doi.org/10.1109/TR.2010.2103990 doi: 10.1109/TR.2010.2103990
|
| [113] |
A. A. Dror, N. Morozov, A. Daoud, Y. Namir, O. Yakir, Y. Shachar, et al., Pre-infection 25-hydroxyvitamin D3 levels and association with severity of COVID-19 illness, PLoS One, 17 (2022), e0263069. https://doi.org/10.1371/journal.pone.0263069 doi: 10.1371/journal.pone.0263069
|
| [114] |
J. Ng, Y. R. Stovezky, D. J. Brenner, S. C. Formenti, I. Shuryak, Development of a model to estimate the association between delay in cancer treatment and local tumor control and risk of metastates, JAMA Netw. Open, 4 (2021), 1-10. https://doi.org/10.1001/jamanetworkopen.2020.34065 doi: 10.1001/jamanetworkopen.2020.34065
|
| [115] |
A. Talkington, R. Durrett, Estimating tumor growth rates in vivo, Bull. Math. Biol., 77 (2015), 1934-1954. https://doi.org/10.1007/s11538-015-0110-8 doi: 10.1007/s11538-015-0110-8
|
| [116] |
C. Vaghi, A. Rodallec, R. Fanciullino, J. Ciccolini, J. P. Mochel, M. Mastri, Population modeling of tumor growth curves and the reduced Gompertz model improve prediction of the age of experimental tumors, PLoS Comput. Biol., 16 (2020), e1007178. https://doi.org/10.1371/journal.pcbi.1007178 doi: 10.1371/journal.pcbi.1007178
|
| [117] |
D. J. Yin, A. R. Moes, J. G. C. van Hasselt, J. J. Swen, H. J. Guchelaar, A review of mathematical models for tumor dynamics and treatment resistance evolution of solid tumors, CPTPharmacomet. Syst. Pharmacol., 8 (2019), 720-737. https://doi.org/10.1002/psp4.12450 doi: 10.1002/psp4.12450
|
| [118] |
T. L. Jackson, H. M. A. Byrne, A mathematical model to study the effects of drug resistance and vasculature on the response of solid tumors to chemotherapy, Math. Biosci., 104 (2000), 17-38. https://doi.org/10.1016/S0025-5564(99)00062-0 doi: 10.1016/S0025-5564(99)00062-0
|
| [119] |
C. E. Meacham, S. J. Morrison, Tumour heterogeneity and cancer cell plasticity, Nature, 501 (2013), 328-337. https://doi.org/10.1038/nature12624 doi: 10.1038/nature12624
|
| [120] |
X. Sun, J. Bao, Y. Z. Shao, Mathematical modeling of therapy-induced cancer drug resistance: Connecting cancer mechanisms to population survival rates, Sci. Rep., 6 (2016) 22498. https://doi.org/10.1038/srep22498 doi: 10.1038/srep22498
|
| [121] |
K. Taniguchi, J. Okami, K. Kodama, M. Higashiyama, K. Kato, Intratumor heterogeneity of epidermal growth factor receptor mutations in lung cancer and its correlation to the response to gefitinib, Cancer Sci., 99 (2008), 929-935. https://doi.org/10.1111/j.1349-7006.2008.00782.x doi: 10.1111/j.1349-7006.2008.00782.x
|
| [122] |
E. R. Lestari, H. Arifah, Dynamics of a mathematical model of cancer cells with chemotherapy, J. Phys. Conf. Ser., 1320 (2019), 1-8. https://doi.org/10.1088/1742-6596/1320/1/012026 doi: 10.1088/1742-6596/1320/1/012026
|
| [123] |
H. Pham, A dynamic model of multiple time-delay interactions between the virus-infected cells and body's immune system with autoimmune diseases, Axioms, 10 (2021), 216. https://doi.org/10.3390/axioms10030216 doi: 10.3390/axioms10030216
|
| [124] |
H. Pham, Mathematical modeling the time-delay interactions between tumor viruses and the immune system with the effects of chemotherapy and autoimmune Diseases, Mathematics, 10 (2022), 756. https://doi.org/10.3390/math10050756 doi: 10.3390/math10050756
|
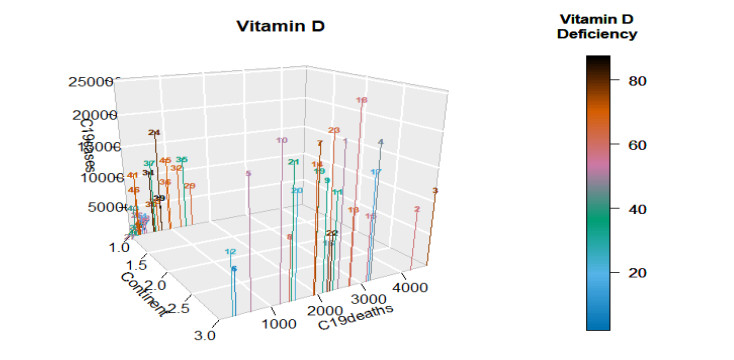
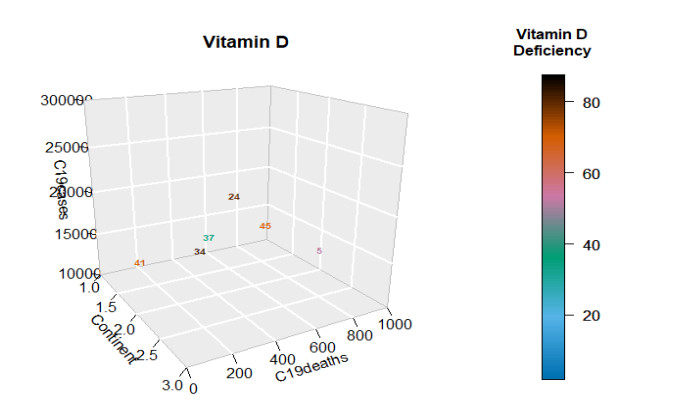
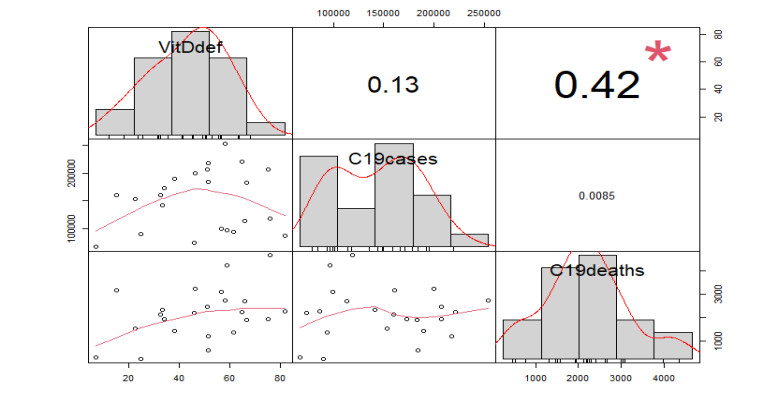
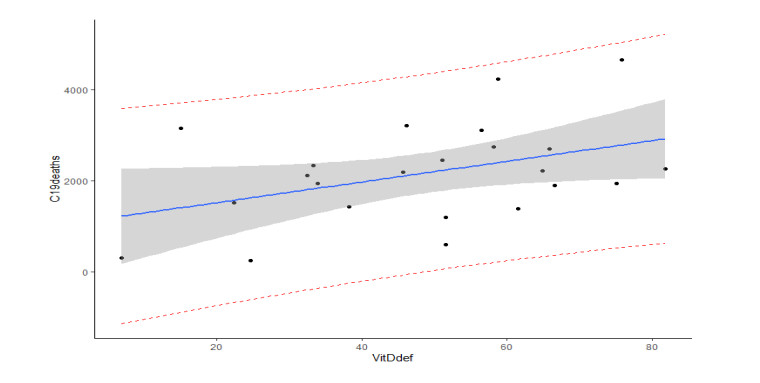
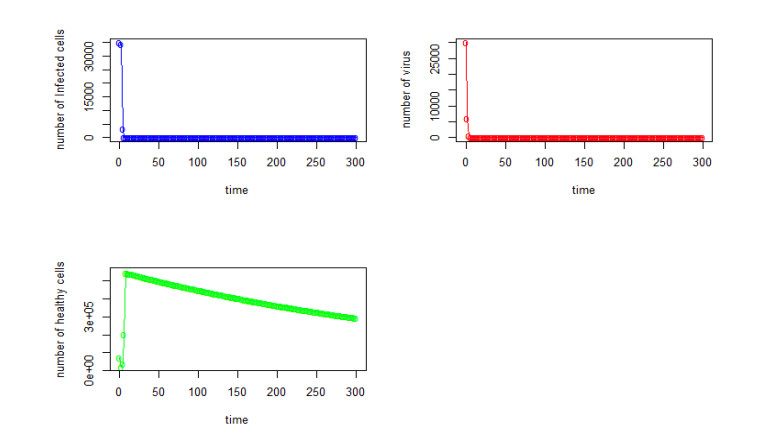
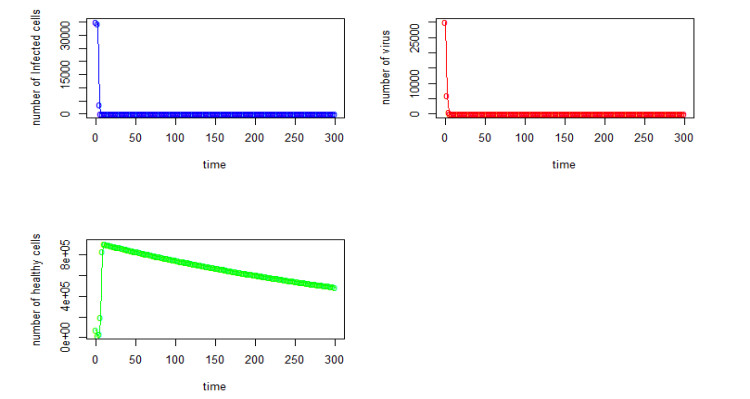
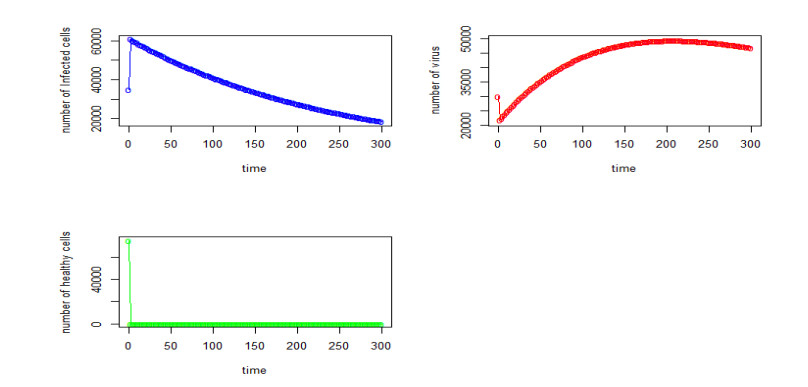
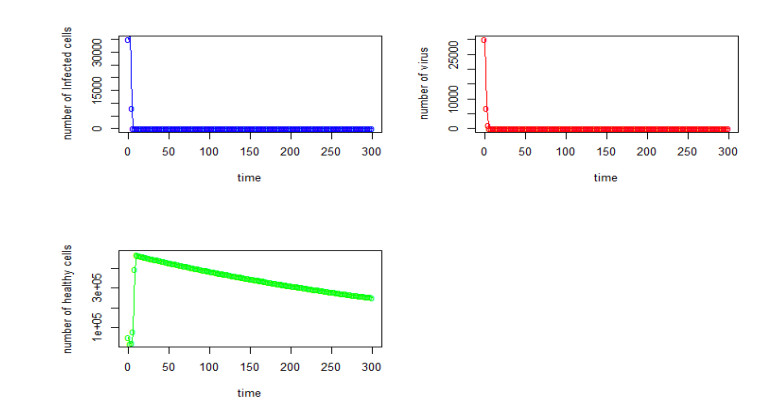
Figures(22) / Tables(4)

Hoang Pham. Analyzing the relationship between the vitamin D deficiency and COVID-19 mortality rate and modeling the time-delay interactions between body's immune healthy cells, infected cells, and virus particles with the effect of vitamin D levels[J]. Mathematical Biosciences and Engineering, 2022, 19(9): 8975-9004. doi: 10.3934/mbe.2022417







 DownLoad:
DownLoad: