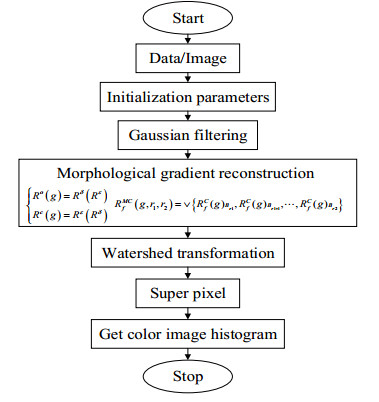
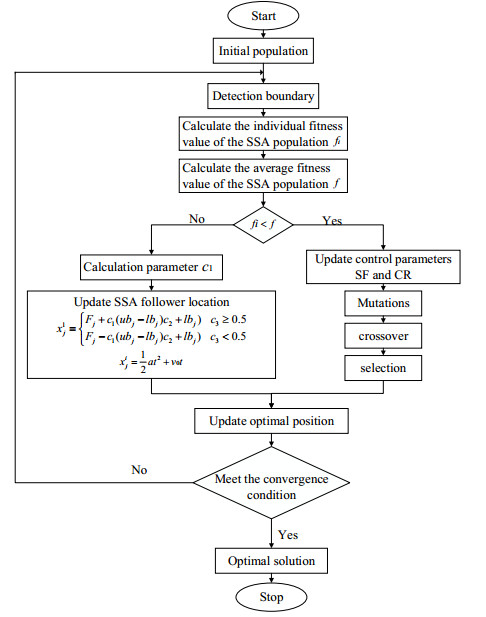
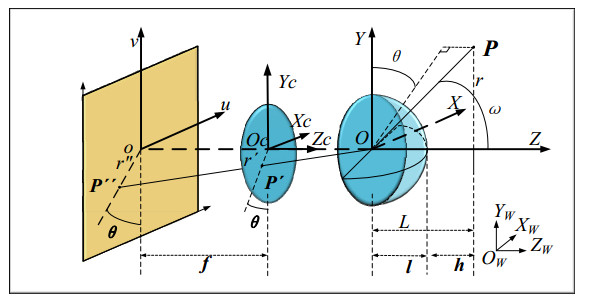
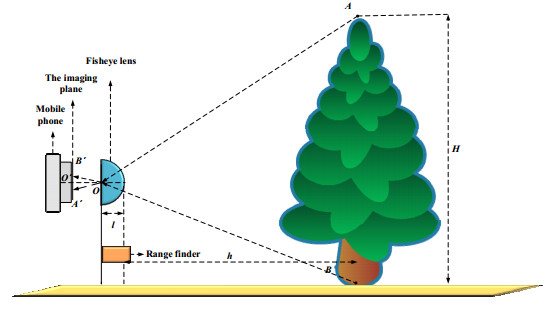
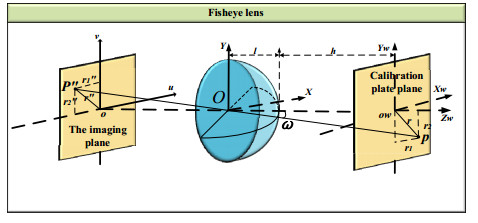
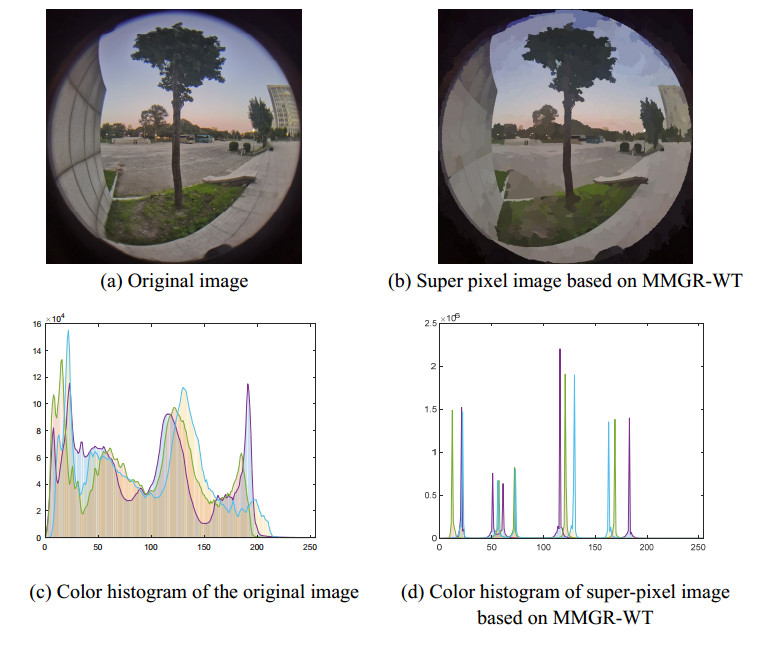
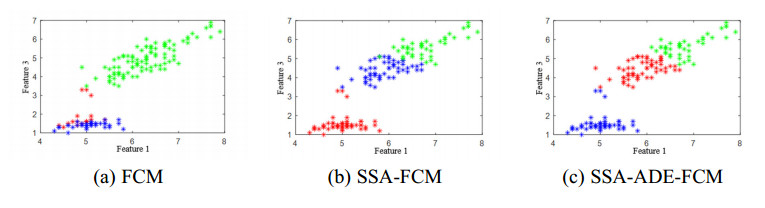
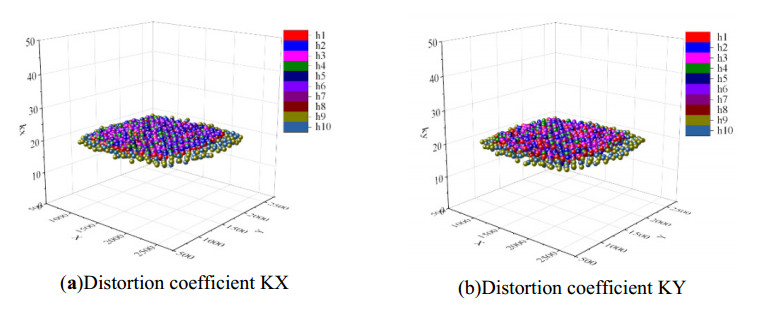
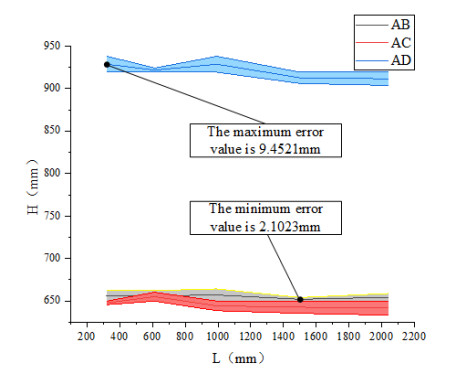
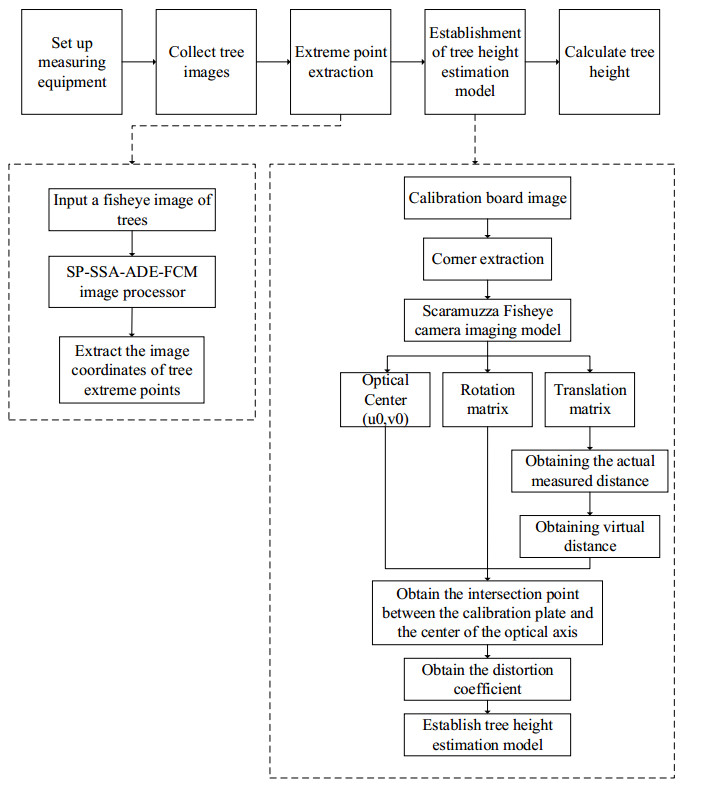
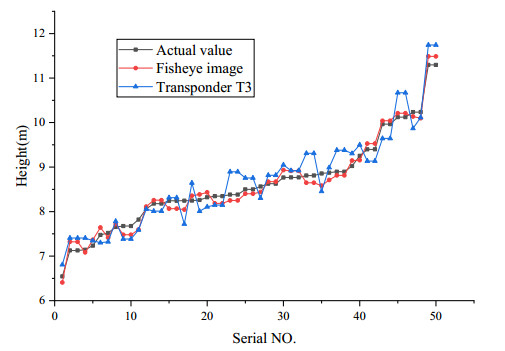
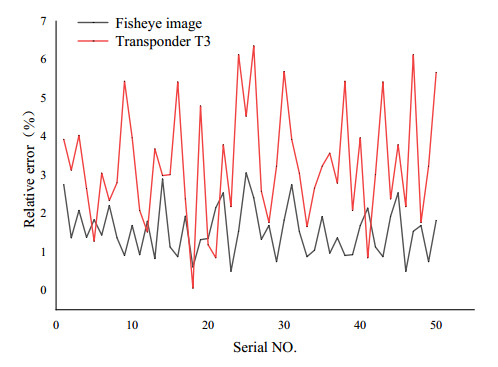
The height of standing trees is an important index in forestry research. This index is not only hard to measure directly but also the environmental factors increase the measurement difficulty. Therefore, the measurement of the height of standing trees is always a problem that experts and scholars are trying to improve. In this study, improve fuzzy c-means algorithm to reduce the calculation time and improve the clustering effect, used on this image segmentation technology, a highly robust non-contact measuring method for the height of standing trees was proposed which is based on a smartphone with a fisheye lens. While ensuring the measurement accuracy, the measurement stability is improved. This method is simple to operate, just need to take a picture of the standing tree and determine the shooting distance to complete the measurement. The purpose of the fisheye lens is to ensure that the tree remains intact in the photograph and to reduce the shooting distance. The results of different stability experiments show that the measurement error ranged from -0.196m to 0.195m, and the highest relative error of tree measurement was 3.05%, and the average relative error was 1.45%. Analysis shows that compared with previous research, this method performs better at all stages. The proposed approach can provide a new way to obtain tree height, which can be used to analyze growing status and change in contrast height because of high accuracy and permanent preservation of images.
Citation: Jiayin Song, Yue Zhao, Zhixiang Chi, Qiang Ma, Tianrui Yin, Xiaopeng Zhang. Improved FCM algorithm for fisheye image cluster analysis for tree height calculation[J]. Mathematical Biosciences and Engineering, 2021, 18(6): 7806-7836. doi: 10.3934/mbe.2021388
The height of standing trees is an important index in forestry research. This index is not only hard to measure directly but also the environmental factors increase the measurement difficulty. Therefore, the measurement of the height of standing trees is always a problem that experts and scholars are trying to improve. In this study, improve fuzzy c-means algorithm to reduce the calculation time and improve the clustering effect, used on this image segmentation technology, a highly robust non-contact measuring method for the height of standing trees was proposed which is based on a smartphone with a fisheye lens. While ensuring the measurement accuracy, the measurement stability is improved. This method is simple to operate, just need to take a picture of the standing tree and determine the shooting distance to complete the measurement. The purpose of the fisheye lens is to ensure that the tree remains intact in the photograph and to reduce the shooting distance. The results of different stability experiments show that the measurement error ranged from -0.196m to 0.195m, and the highest relative error of tree measurement was 3.05%, and the average relative error was 1.45%. Analysis shows that compared with previous research, this method performs better at all stages. The proposed approach can provide a new way to obtain tree height, which can be used to analyze growing status and change in contrast height because of high accuracy and permanent preservation of images.

| [1] |
Y. Wang, M. Lehtomäki, X. Liang, J. Pyörälä, A. Kukko, A. Jaakkola, et al., Is field-measured tree height as reliable as believed a comparison study of tree height estimates from field measurement, airborne laser scanning and terrestrial laser scanning in a boreal forest, ISPRS J. Photogramm., 147 (2019), 132-145. doi: 10.1016/j.isprsjprs.2018.11.008

|
| [2] | I.S. Saliu, B. Satyanarayana, M.A. B. Fisol, G. Wolswijk, C. Decannière, R. Lucas, et al., An accuracy analysis of mangrove tree height mensuration using forestry techniques, hypsometers and UAVs, Estuar. Coast. Shelf Sci., 248 (2020), 106971. |
| [3] |
D.W. Wanik, J.R. Parent, E.N. Anagnostou, B.M. Hartman, Using vegetation management and LiDAR-derived tree height data to improve outage predictions for electric utilities, Electr. Pow. Syst. Res., 146 (2017), 236-245. doi: 10.1016/j.epsr.2017.01.039
|
| [4] |
Y. D. Huang, M. Z. Li, S. Q. Ren, M. J. Wang, P. Y. Cui, Impacts of tree-planting pattern and trunk height on the airflow and pollutant dispersion inside a street canyon, Build. Environ., 165 (2019), 106385. doi: 10.1016/j.buildenv.2019.106385
|
| [5] |
Y. Xu, C. Li, Z. Sun, L. Jiang, J. Fang, Tree height explains stand volume of closed-canopy stands: evidence from forest inventory data of China, Forest Ecol. Manag., 438 (2019), 51-56. doi: 10.1016/j.foreco.2019.01.054
|
| [6] |
C. Cabo, C. Ordóñez, C. A. López-Sánchez, J. Armesto, Automatic dendrometry: tree detection, tree height and diameter estimation using terrestrial laser scanning, Int. J. Appl. Earth Obs., 69 (2018), 164-174. doi: 10.1016/j.jag.2018.01.011
|
| [7] | J. S. Cui, J. Huo, M. Yang, Y. K. Wang, Research on the rigid body posture measurement using monocular vision by coplanar feature points, Optics, 126 (2015), 5423-5429. |
| [8] |
H. Du, M. G. Li, The study for particle image velocimetry system based on binocular vision, Measurement, 42 (2009), 619-627. doi: 10.1016/j.measurement.2008.10.011
|
| [9] |
X. Hu, H. Zheng, Y. Chen, L. Chen, Dense crowd counting based on perspective weight model using a fisheye camera, Optik, 126 (2015), 123-130. doi: 10.1016/j.ijleo.2014.08.132
|
| [10] |
H. Kim, J. Jung, L. Paik, Fisheye lens camera based surveillance system for wide field of view monitoring, Optik, 127 (2016), 5636-5646. doi: 10.1016/j.ijleo.2016.03.069
|
| [11] |
J. Zhu, J. Zhu, X. Wan, C. Wu, C. Xu, Object detection and localization in 3D environment by fusing raw fisheye image and attitude data, J. Vis. Commun. Image R., 59 (2019), 128-139. doi: 10.1016/j.jvcir.2019.01.005
|
| [12] |
W. C. Wang, C. H. Hwang, C. I. Chu, Y. H. Chen, Displacement measurement of interior wall of hollow cylinder by digital image correlation method using fisheye lens, Procedia Eng., 79 (2014), 437-446. doi: 10.1016/j.proeng.2014.06.365
|
| [13] | Z. Y. Liu, F. Ding, Y. Xu, X. Han, Background dominant colors extraction method based on color image quick fuzzy c-means clustering algorithm, Def. Technol., (2020). |
| [14] | J. C. Dunn, A fuzzy relative of the ISODATA process and its use in detecting compact well-separated clusters, Taylor Francis, 3 (1973), 32-57. |
| [15] | Y. H. Chen, L. J. Zhang, H. T. Lang, Acceleration of fuzzy c-means clustering based target detection for large size image, J. Ocean U. China, 47 (2017), 94-100. |
| [16] | S. Sandhya, B. Chidambararajan, M. S. Kumar, FCM-based segmentation and neural network classification of tumor in brain mri images, in Intelligence in Big Data Technologies-Beyond the Hype, Springer, (2021), 371-378. |
| [17] |
B. J. Shankar, K. Murugan, A. Obulesu, D. Finney, R. Anitha, MRI image segmentation using bat optimization algorithm with fuzzy c means (BOA-FCM) clustering, J. Med. Imag. Health Inform., 11 (2021), 661-666. doi: 10.1166/jmihi.2021.3365
|
| [18] |
M. Li, Y. Wang, Q. Sun, Y. Liu, Research of ASW-FCM-based algorithm for clustered wind turbine group equivalent modeling, J. Electr. Eng. Technol., 15 (2020), 1555-1566. doi: 10.1007/s42835-020-00439-0
|
| [19] | J. Fan, J. Wang, A two-phase fuzzy clustering algorithm based on neurodynamic optimization with its application for polsar image segmentation, IEEE Trans. Fuzzy Syst., 26 (2016), 72-83. |
| [20] | R. Shang, P. Tian, L. Jiao, R. Stolkin, J. Feng, B. Hou, et al., A spatial fuzzy clustering algorithm with kernel metric based on immune clone for SAR image segmentation, IEEE J. Stars., 9 (2016), 1640-1652. |
| [21] |
Y. Zhang, X. Bai, R. Fan, Z. Wang, Deviation-sparse fuzzy c-means with neighbor information constraint, IEEE Trans. Fuzzy Syst., 27 (2019), 185-199. doi: 10.1109/TFUZZ.2018.2883033
|
| [22] |
S. Krinidis, V. Chatzis, A robust fuzzy local information C-means clustering algorithm, IEEE Trans. Image Process., 19 (2010), 1328-1337. doi: 10.1109/TIP.2010.2040763
|
| [23] | X. Bai, Z. Chen, Y. Zhang, Z. Liu, Y. Lu, Infrared ship target segmentation based on spatial information improved FCM, IEEE Trans. Cyber., 46 (2015), 3259-3271. |
| [24] |
H. Zhang, Q. Wang, W. Shi, M. Hao, A novel adaptive fuzzy local information C-Means clustering algorithm for remotely sensed imagery classification, IEEE Trans. Geosci. Remote Sen., 55 (2017), 5057-5068. doi: 10.1109/TGRS.2017.2702061
|
| [25] | P. K. Mishro, S. Agrawal, L. Dora, R. Panda, A fuzzy C-means clustering approach to HMRF-EM model for MRI brain tissue segmentation, in 2017 6th International Conference on Computer Applications in Electrical Engineering-Recent Advances (CERA), Springer, (2017), 371-376. |
| [26] |
H. Zhang, Q. Wu, Y. Zheng, T. M. Nguyen, D. Wang, Effective fuzzy clustering algorithm with Bayesian model and mean template for image segmentation, IET Image Process, 8 (2014), 571-581. doi: 10.1049/iet-ipr.2013.0178
|
| [27] | L. Szilagyi, Z. Benyo, S. M. Szilagyi, H. S. Adam, MR brain image segmentation using an enhanced fuzzy C-means algorithm, in Proceedings of International Conference of the IEEE Engineering in Medicine & Biology Society, Springer, (2003), 724-726. |
| [28] |
D. Comaniciu, P. Meer, Mean shift: a robust approach toward feature space analysis, IEEE Transactions Pattern Anal. Mach. Intell., 24 (2002), 603-619. doi: 10.1109/34.1000236
|
| [29] | A. Y. Ng, M. I. Jordan, Y. Weiss, On spectral clustering: analysis and an algorithm, in Advances in Neural Information Processing Systems, Springer, (2002), 849-856. |
| [30] | T. Lei, X. Jia, Y. Zhang, S. Liu, H. Meng, A.K. Nandi, Superpixel-based fast fuzzy c-means clustering for color image segmentation, IEEE Trans. Fuzzy Syst., (2018), 1753-1766. |
| [31] |
P. Arbeláez, M. Maire, C. Fowlkes, J. Malik, Contour detection and hierarchical image segmentation, IEEE Transactions Pattern Anal. Mach. Intell., 33 (2011), 898-916. doi: 10.1109/TPAMI.2010.161
|
| [32] |
Q. Lin, Q. Zhu, P. Huang, J. Chen, Z. Ming, J. Yu, Computers, A novel hybrid multi-objective immune algorithm with adaptive differential evolution, Comput. Oper. Res., 62 (2015), 95-111. doi: 10.1016/j.cor.2015.04.003
|
| [33] | L. Zuo, C. Luo, Y. Zuo, Paralleled segmentation cluster algorithm based on En FCM for large-scale image, Microcomput Its Appl., 34 (2015), 55-58. |
| [34] |
J. Song, Z. Zhang, A modified robust fcm model with spatial constraints for brain MR image segmentation, Information, 10 (2019), 74. doi: 10.3390/info10020074
|
| [35] |
H. X. Pei, Z. R. Zheng, C. Wang, C. N. Li, Y. H. Yao, D-fcm: density based fuzzy C-means clustering algorithm with application in medical image segmentation, Procedia Comput. Sci., 122 (2017), 407-414. doi: 10.1016/j.procs.2017.11.387
|






Figures(18) / Tables(7)

Jiayin Song, Yue Zhao, Zhixiang Chi, Qiang Ma, Tianrui Yin, Xiaopeng Zhang. Improved FCM algorithm for fisheye image cluster analysis for tree height calculation[J]. Mathematical Biosciences and Engineering, 2021, 18(6): 7806-7836. doi: 10.3934/mbe.2021388







 DownLoad:
DownLoad: