Citation: Faisal Mehmood Butt, Lal Hussain, Anzar Mahmood, Kashif Javed Lone. Artificial Intelligence based accurately load forecasting system to forecast short and medium-term load demands[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 400-425. doi: 10.3934/mbe.2021022

| [1] | Y. Al-Rashid, L. D. Paarmann, Short-term electric load forecasting using neural network models, in: Proc. 39th Midwest Symp. Circuits Syst., IEEE, (1996), 1436–1439. |
| [2] |
T. Hong, P. Pinson, S. Fan, H. Zareipour, A. Troccoli, R. J. Hyndman, Probabilistic energy forecasting: Global Energy Forecasting Competition 2014 and beyond, Int. J. Forecast., 32 (2016), 896–913. doi: 10.1016/j.ijforecast.2016.02.001

|
| [3] |
T. Hong, P. Pinson, S. Fan, Global Energy Forecasting Competition 2012, Int. J. Forecast., 30 (2014), 357–363. doi: 10.1016/j.ijforecast.2013.07.001
|
| [4] | L. Hussain, M.S. Nadeem, S. A. A.Shah, Short term load forecasting system based on Support Vector Kernel methods, Int. J. Comput. Sci. Inf. Technol., 6 (2014), 93–102. |
| [5] |
G. Gross, F. D. Galiana, Short-term load forecasting, Proc. IEEE., 75 (1987), 1558–1573. doi: 10.1109/PROC.1987.13927
|
| [6] | P. J. Santos, A. G. Martins, A. J. Pires, Short-term load forecasting based on ANN applied to electrical distribution substations, in: 39th Int. Univ. Power Eng. Conf. UPEC 2004 - Conf. Proc., IEEE, 1(2004), 427–432. |
| [7] |
E. Hossain, I. Khan, F. Un-Noor, S. S. Sikander, M. S. H. Sunny, Application of big data and machine learning in smart grid, and associated security concerns: A Review, IEEE Access, 7 (2019), 13960–13988. doi: 10.1109/ACCESS.2019.2894819
|
| [8] |
H. J. Sadaei, P. C. D.L. e Silva, F. G. Guimarães, M. H. Lee, Short-term load forecasting by using a combined method of convolutional neural networks and fuzzy time series, Energy, 175 (2019), 365–377. doi: 10.1016/j.energy.2019.03.081
|
| [9] |
H. M. Al-Hamadi, S. A. Soliman, Short-term electric load forecasting based on Kalman filtering algorithm with moving window weather and load model, Electr. Power Syst. Res., 68 (2004), 47–59. doi: 10.1016/S0378-7796(03)00150-0
|
| [10] |
K.-B. Song, Y.-S. Baek, D. H. Hong, G. Jang, Short-term load forecasting for the holidays using fuzzy linear regression method, IEEE Trans. Power Syst., 20 (2005), 96–101. doi: 10.1109/TPWRS.2004.835632
|
| [11] |
D. K. Ranaweera, N. F. Hubele, G. G. Karady, Fuzzy logic for short term load forecasting, Int. J. Electr. Power Energy Syst., 18 (1996), 215–222. doi: 10.1016/0142-0615(95)00060-7
|
| [12] |
Y. He, Q. Xu, J. Wan, S. Yang, Short-term power load probability density forecasting based on quantile regression neural network and triangle kernel function, Energy, 114 (2016), 498–512. doi: 10.1016/j.energy.2016.08.023
|
| [13] |
M. Q. Raza, A. Khosravi, A review on artificial intelligence based load demand forecasting techniques for smart grid and buildings, Renew. Sustain. Energy Rev., 50 (2015), 1352–1372. doi: 10.1016/j.rser.2015.04.065
|
| [14] | L. C. P. Velasco, C. R. Villezas, P. N. C. Palahang, J. A. A. Dagaang, Next day electric load forecasting using Artificial Neural Networks, in: 2015 Int. Conf. Humanoid, Nanotechnology, Inf. Technol. Control. Environ. Manag., IEEE, 2015, 1–6. |
| [15] |
L. Hernández, C. Baladrón, J. M. Aguiar, L. Calavia, B. Carro, A. Sánchez-Esguevillas, et al., Artificial Neural Network for short-term load forecasting in distribution systems, Energies, 7 (2014), 1576–1598. doi: 10.3390/en7031576
|
| [16] |
J. Buitrago, S. Asfour, Short-term forecasting of electric loads using nonlinear autoregressive Artificial Neural Networks with exogenous vector inputs, Energies, 10 (2017), 40. doi: 10.3390/en10010040
|
| [17] |
L. Suganthi, S. Iniyan, A. A. Samuel, Applications of fuzzy logic in renewable energy systems- A review, Renew. Sustain. Energy Rev., 48 (2015), 585–607. doi: 10.1016/j.rser.2015.04.037
|
| [18] |
K-H. Kim, J-K. Park, K-J. Hwang, S-H. Kim, Implementation of hybrid short-term load forecasting system using artificial neural networks and fuzzy expert systems, IEEE Trans. Power Syst., 10 (1995), 1534–1539. doi: 10.1109/59.466492
|
| [19] |
D. X. Niu, H. F. Shi, D. D. Wu, Short-term load forecasting using bayesian neural networks learned by Hybrid Monte Carlo algorithm, Appl. Soft Comput., 12 (2012), 1822–1827. doi: 10.1016/j.asoc.2011.07.001
|
| [20] |
F. Kaytez, M. C. Taplamacioglu, E. Cam, F. Hardalac, Forecasting electricity consumption: A comparison of regression analysis, neural networks and least squares support vector machines, Int. J. Electr. Power Energy Syst., 67 (2015), 431–438. doi: 10.1016/j.ijepes.2014.12.036
|
| [21] | X. Li, X. Wu, Constructing long short-term memory based deep recurrent neural networks for large vocabulary speech recognition, in: 2015 IEEE Int. Conf. Acoust. Speech Signal Process, IEEE, 2015, 4520–4524. |
| [22] |
R. J. Williams, D. Zipser, A learning algorithm for continually running fully recurrent Neural Networks, Neural Comput., 1 (1989), 270–280. doi: 10.1162/neco.1989.1.2.270
|
| [23] |
M. Ghofrani, M. Ghayekhloo, A. Arabali, A. Ghayekhloo, A hybrid short-term load forecasting with a new input selection framework, Energy, 81 (2015), 777–786. doi: 10.1016/j.energy.2015.01.028
|
| [24] |
G.-F. Fan, L.-L. Peng, W.-C. Hong, Short term load forecasting based on phase space reconstruction algorithm and bi-square kernel regression model, Appl. Energy, 224 (2018), 13–33. doi: 10.1016/j.apenergy.2018.04.075
|
| [25] |
R. Mamlook, O. Badran, E. Abdulhadi, A fuzzy inference model for short-term load forecasting, Energy Policy, 37 (2009), 1239–1248. doi: 10.1016/j.enpol.2008.10.051
|
| [26] |
K. Metaxiotis, A. Kagiannas, D. Askounis, J. Psarras, Artificial intelligence in short term electric load forecasting: a state-of-the-art survey for the researcher, Energy Convers. Manag., 44 (2003), 1525–1534. doi: 10.1016/S0196-8904(02)00148-6
|
| [27] | K. Fukushima, S. Miyake, A self-organizing neural network model for a mechanism of visual pattern recognition, in: Comp. cooperation Neural Net., Springer Berlin Heidelb., 1982,267–85. |
| [28] | Y. LeCun, P. Haffner, L. Bottou, Y. Bengio, Object recognition with gradient-based learning, in: Shape, Cont. grop. comp. vision., Springer Berlin Heidelb., 1999,319–345. |
| [29] |
Y. Guo, Y. Liu, A. Oerlemans, S. Lao, S. Wu, Y. Guo, et al., Deep learning for visual understanding: A review, Neurocomputing, 187 (2016), 27–48. doi: 10.1016/j.neucom.2015.09.116
|
| [30] |
H.-K. Yu, Weighted fuzzy time series models for TAIEX forecasting, Phys. A Stat. Mech. Appl., 349 (2005), 609–624. doi: 10.1016/j.physa.2004.11.006
|
| [31] |
N.-Y. Wang, S.-M. Chen, Temperature prediction and TAIFEX forecasting based on automatic clustering techniques and two-factors high-order fuzzy time series, Expert Syst. Appl., 36 (2009), 2143–2154. doi: 10.1016/j.eswa.2007.12.013
|
| [32] |
W.-J. Lee, J. Hong, A hybrid dynamic and fuzzy time series model for mid-term power load forecasting, Int. J. Electr. Power Energy Syst., 64 (2015), 1057–1062. doi: 10.1016/j.ijepes.2014.08.006
|
| [33] |
R. Efendi, Z. Ismail, M. M. Deris, A new linguistic out-sample approach of fuzzy time series for daily forecasting of Malaysian electricity load demand, Appl. Soft Comput., 28 (2015), 422–430. doi: 10.1016/j.asoc.2014.11.043
|
| [34] |
R. Enayatifar, H. J. Sadaei, A. H. Abdullah, A. Gani, Imperialist competitive algorithm combined with refined high-order weighted fuzzy time series (RHWFTS–ICA) for short term load forecasting, Energy Convers. Manag., 76 (2013), 1104–1116. doi: 10.1016/j.enconman.2013.08.039
|
| [35] |
L. Hussain, W. Aziz, J. S. Alowibdi, N. Habib, M. Rafique, S. Saeed, et al., Symbolic time series analysis of electroencephalographic (EEG) epileptic seizure and brain dynamics with eye-open and eye-closed subjects during resting states, J. Physiol. Anthropol., 36 (2017), 21. doi: 10.1186/s40101-017-0136-8
|
| [36] |
L. Hussain, Detecting epileptic seizure with different feature extracting strategies using robust machine learning classification techniques by applying advance parameter optimization approach, Cogn. Neurodyn., 12 (2018), 271–294. doi: 10.1007/s11571-018-9477-1
|
| [37] | L. Hussain, I. A. Awan, W. Aziz, S. Saeed, A. Ali, F. Zeeshan, et al., Detecting congestive heart failure by extracting multimodal features and employing machine learning techniques, Biomed. Res. Int., 2020 (2020), 1–19. |
| [38] |
Y. Asim, B. Raza, A. Kamran, M. Saima, A. K. Malik, S. Rathore, et al., A multi-modal, multi-atlas-based approach for Alzheimer detection via machine learning, Int. J. Imag. Syst. Tech., 28 (2018), 113–123. doi: 10.1002/ima.22263
|
| [39] |
L. Hussain, S. Saeed, I. A. Awan, A. Idris, M. S. A. Nadeem, Q.-A. Chaudhry, et al., Detecting brain tumor using machines learning techniques based on different features extracting strategies, Curr. Med. Imag. Rev., 15 (2019), 595–606. doi: 10.2174/1573405614666180718123533
|
| [40] |
L. Hussain, W. Aziz, S. Saeed, I. A. Awan, A. A. Abbasi, N. Maroof, Arrhythmia detection by extracting hybrid features based on refined Fuzzy entropy (FuzEn) approach and employing machine learning techniques, Waves Rand. Complex Media, 30 (2020), 656–686. doi: 10.1080/17455030.2018.1554926
|
| [41] |
L. Hussain, W. Aziz, A. A. Alshdadi, M. S. A. Nadeem, I. R. Khan, Q.-A. Chaudhry, Analyzing the dynamics of lung cancer imaging data using refined fuzzy entropy methods by extracting different features, IEEE Access, 7 (2019), 64704–64721. doi: 10.1109/ACCESS.2019.2917303
|
| [42] | A. A. Abbasi, L. Hussain, I. A. Awan, I. Abbasi, A. Majid, M. S. A. Nadeem, Q. -A. Chaudhary, Detecting prostate cancer using deep learning convolution neural network with transfer learning approach, Cogn. Neurodyn., 5 (2020), 1–11. |
| [43] |
L. Hussain, A. Ahmed, S. Saeed, S. Rathore, I. A. Awan, S. A. Shah, et al., Prostate cancer detection using machine learning techniques by employing combination of features extracting strategies, Cancer Biomark., 21 (2018), 393–413. doi: 10.3233/CBM-170643
|
| [44] | C. Lang, F. Steinborn, O. Steffens, E. W. Lang, Applying a 1D-CNN network to electricity load forecasting, in: Int. conf. time series forecas., Springer, Cham, 2020,205–218. |
| [45] | N. Javaid, NADEEM: A Novel Reliable Data Delivery Routing Protocol for Underwater WSNs, in: Works. Int. Conf. Adv. Inf. Netw. App., Springer, Cham, 2019,103–115. |
| [46] |
D. Hussain, T. Hussain, A. A. Khan, S. A. A. Naqvi, A. Jamil, A deep learning approach for hydrological time-series prediction: A case study of Gilgit river basin, Earth Sci. Inform., 13 (2020), 915–927. doi: 10.1007/s12145-020-00477-2
|
| [47] | A.M. Tudose, D.O. Sidea, I. I. Picioroaga, V. A. Boicea, C. Bulac, A CNN Based Model for Short-Term Load Forecasting: A Real Case Study on the Romanian Power System, in: 2020 55th Int. Univ. Power Eng. Conf., IEEE, 2020, 1–6. |
| [48] | M. Solas, N. Cepeda, J. L. Viegas, Convolutional Neural Network for Short-term Wind Power Forecasting, in: 2019 IEEE PES Innov. Smart Grid Technol. Eur., IEEE, 2019, 1–5. |
| [49] |
A. E. Khantach, M. Hamlich, N. E. Belbounaguia, Short-term load forecasting using machine learning and periodicity decomposition, AIMS Energy, 7 (2019), 382–394. doi: 10.3934/energy.2019.3.382
|
| [50] | G. Dudek, Forecasting time series with multiple seasonal cycles using neural networks with local learning, in: Int. Conf. Artif. Intell. Soft Comput., Springer, Berlin, Heidelberg., 2013, 52–63. |
| [51] |
M. A. Ghorbani, R. C. Deo, Z. M. Yaseen, M. H. Kashani, B. Mohammadi, Pan evaporation prediction using a hybrid multilayer perceptron-firefly algorithm (MLP-FFA) model: Case study in North Iran, Theoret. App. Climat., 133 (2018), 1119–1131. doi: 10.1007/s00704-017-2244-0
|
| [52] | M. E. Günay, Forecasting annual gross electricity demand by artificial neural networks using predicted values of socio-economic indicators and climatic conditions: Case of Turkey, Energy Policy, 90 (2016), 92–101. |
| [53] |
G. Dudek, Multilayer perceptron for short-term load forecasting : From global to local approach, Neural Comput. Appl., 32 (2020), 3695–3707. doi: 10.1007/s00521-019-04130-y
|
| [54] | X. Glorot, Y. Bengio, Understanding the difficulty of training deep feedforward neural networks, in: Proc. thirteenth int. conf. artif. intell. stat., 2010,249–256. |
| [55] |
S. Hochreiter, J Schmidhuber, Long short-term memory, Neural Comput., 9 (1997), 1735–1780. doi: 10.1162/neco.1997.9.8.1735
|
| [56] |
H. Zheng, J. Yuan, L. Chen, Short-term load forecasting using EMD-LSTM neural networks with a Xgboost algorithm for feature importance evaluation, Energies, 10 (2017), 1168. doi: 10.3390/en10081168
|
| [57] | Y. Zhao, Y. Shen, Y. Zhu, J. Yai, Forecasting wavelet transformed time series with attentive neural networks, in: 2018 IEEE Int. Conf. Data Min., IEEE, 2018, 1452–1457. |
| [58] | K. Greff, R. K. Srivastava, J. Koutnik, B. R. Steunebrink, J. Schmidhuber, LSTM: A Search Space Odyssey, IEEE Trans. Neural Networks Learn. Syst., 28 (2016), 2222–2232. |
| [59] |
P. Nagabushanam, S. T. George, S. Radha, EEG signal classification using LSTM and improved neural network algorithms, Soft Comput., 24 (2020), 9981–10003. doi: 10.1007/s00500-019-04515-0
|
| [60] |
V. Y. Senyurek, M. H. Imtiaz, P. Belsare, S. Tiffany, E. Sazonov, A CNN-LSTM neural network for recognition of puffing in smoking episodes using wearable sensors, Biomed. Eng. Lett., 10 (2020), 195–203. doi: 10.1007/s13534-020-00147-8
|
| [61] |
A. Graves, J. Schmidhuber, Framewise phoneme classification with bidirectional LSTM and other neural network architectures, Neural Networks, 18 (2005), 602–610. doi: 10.1016/j.neunet.2005.06.042
|
| [62] | A. Graves, Long Short-Term Memory, in: Sup. Seq. Label.Recurr. Neural Networks. Stud. Comput. Intell., Springer Berlin Heidelberg, 2012, 37–45. |
| [63] |
X. Ma, Z. Tao, Z. Wang, H. Yu, Y. Wang, Long short-term memory neural network for traffic speed prediction using remote microwave sensor data, Transp. Res. Part C Emerg. Technol., 54 (2015), 187–197. doi: 10.1016/j.trc.2015.03.014
|
| [64] |
E. Shelhamer, J. Long, T. Darrell, Fully convolutional networks for semantic segmentation, IEEE Trans. Pattern Anal. Mach. Intell., 39 (2017), 640–651. doi: 10.1109/TPAMI.2016.2572683
|
| [65] | Y. Taigman, M. Yang, M. Ranzato, L. Wolf, DeepFace: Closing the gap to human-level performance in face verification, in: Proc. IEEE Comput. Soc. Conf. Comput. Vis. Pattern Recognit., 2014, 1701–1708. |
| [66] | A. Karpathy, G. Toderici, S. Shetty, T. Leung, R. Sukthankar, L. Fei-Fei, Large-scale video classification with convolutional neural networks, in: Proc. IEEE Conf. Comput. Vis. Pattern Recognit. IEEE, 2014, 1725–1732. |
| [67] |
T. Young, D. Hazarika, S. Poria, E. Cambria, Recent trends in deep learning based natural language processing, IEEE Comput. Intell. Mag., 13 (2018), 55–75. doi: 10.1109/MCI.2018.2840738
|
| [68] | M. Cai, M. Pipattanasomporn, S. Rahman, Day-ahead building-level load forecasts using deep learning vs. traditional time-series techniques, Appl. Energy, 236 (2019), 1078–1088. |
| [69] |
M. Canizo, I. Triguero, A. Conde, E. Onieva, Multi-head CNN–RNN for multi-time series anomaly detection: An industrial case study, Neurocomputing, 363 (2019), 246–260. doi: 10.1016/j.neucom.2019.07.034
|
| [70] |
Z. Zeng, H. Xiao, X. Zhang, Self CNN-based time series stream forecasting, Electron. Lett., 52 (2016), 1857–1858. doi: 10.1049/el.2016.2626
|
| [71] |
Y. Lecun, L. Bottou, Y. Bengio, P. Haffner, Gradient-based learning applied to document recognition, Proc. IEEE, 86 (1998), 2278–2324. doi: 10.1109/5.726791
|
| [72] | L. Atlas, T. Homma, R. Marks, An artificial neural network for spatio-temporal bipolar patterns: Application to phoneme classification, in: Neural Inf. Process, 1988, 31–40. |
| [73] |
J. Günther, P. M. Pilarski, G. Helfrich, H. Shen, K. Diepold, First steps towards an intelligent laser welding architecture using deep neural networks and reinforcement learning, Procedia Technol., 15 (2014), 474–483. doi: 10.1016/j.protcy.2014.09.007
|
| [74] |
N. Amjady, F. Keynia, Short-term load forecasting of power systems by combination of wavelet transform and neuro-evolutionary algorithm, Energy, 34 (2009), 46–57. doi: 10.1016/j.energy.2008.09.020
|
| [75] | L. Hussain, S. Saeed, A. Idris, I. A. Awan, S. A. Shah, A. Majid, et al., Regression analysis for detecting epileptic seizure with different feature extracting strategies, Biomed. Eng. Biomed. Tech., 64 (2019), 619–642. |
| [76] | S. Du, T. Li, X. Gong, Y. Yang, S.J. Horng, Traffic flow forecasting based on hybrid deep learning framework, in: 2017 12th Int. Conf. Intell. Syst. Knowl. Eng., IEEE, 2017, 1–6. |
| [77] |
P.-H. Kuo, C.-J. Huang, A high precision artificial neural networks model for short-term energy load forecasting, Energies, 11 (2018), 213. doi: 10.3390/en11010213
|
| [78] | G. M. U. Din, A. K. Marnerides, Short term power load forecasting using Deep Neural Networks, in: 2017 Int. Conf. Comput. Netw. Commun., IEEE, (2017), 594–598. |
| [79] |
S. Fan, L. Chen, Short-term load forecasting based on an adaptive hybrid method, IEEE Trans. Power Syst., 21 (2006), 392–401. doi: 10.1109/TPWRS.2005.860944
|
| [80] |
T. Ouyang, Y. He, H. Li, Z. Sun, S. Baek, Modeling and forecasting short-term power load with copula model and deep belief network, IEEE Trans. Emerg. Top. Comput. Intell., 3 (2019), 127–136. doi: 10.1109/TETCI.2018.2880511
|
| [81] | X. Wang, F. Fang, X. Zhang, Y. Liu, L. Wei, Y. Shi, LSTM-based Short-term Load Forecasting for Building Electricity Consumption, in: 2019 IEEE 28th Int. Symp. Ind. Electron., IEEE, 2019, 1418–1423. |
| [82] |
F. Rodrigues, C. Cardeira, J. M. F. Calado, The daily and hourly energy consumption and load forecasting using artificial neural network method: A case study using a set of 93 households in Portugal, Energy Proced., 62 (2014), 220–229. doi: 10.1016/j.egypro.2014.12.383
|
| [83] |
I. Shah, H. Iftikhar, S. Ali, Modeling and forecasting medium-term electricity consumption using component estimation technique, Forecasting, 2 (2020), 163–179. doi: 10.3390/forecast2020009
|
| [84] | S. H. Rafi, N. Al-Masood, Short Term Electric Load Forecasting Using High Precision Convolutional Neural Network, in: 2020 Int. Conf. Comput. Electr. Commun. Eng., IEEE, 2020, 1–7. |
| [85] | S. H. Rafi, N. Al-Masood, Highly Efficient Short Term Load Forecasting Scheme Using Long Short Term Memory Network, in: 2020 8th Int. Electr. Eng. Congr., IEEE, (2020), 1–4. |
| [86] | R. A. Abbasi, N. Javaid, M. N. J. Ghuman, Z. A. Khan, S. Ur Rehman, Amanullah, Short Term Load Forecasting Using XGBoost, in: Work. Int. Conf. Adv. Info. Netw. App., (2019), 1120–1131. |
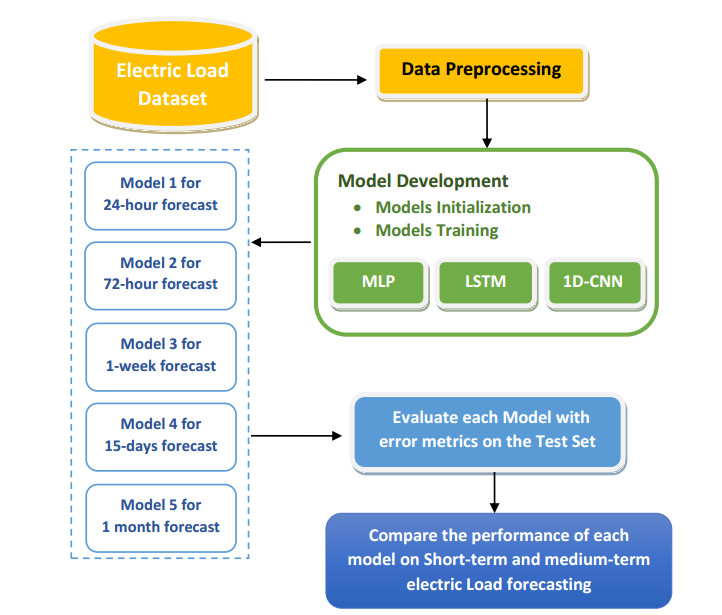
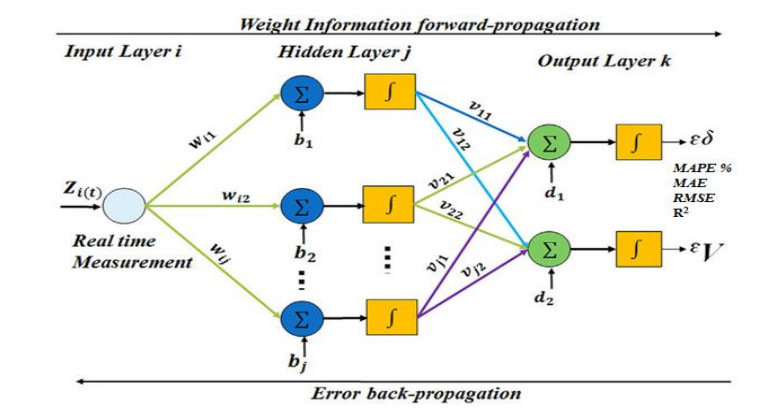
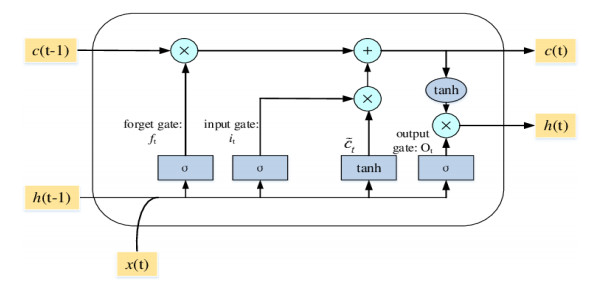
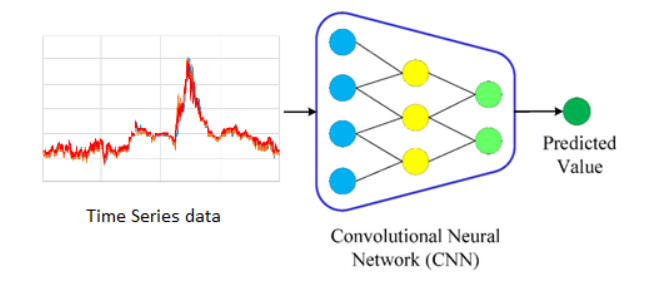
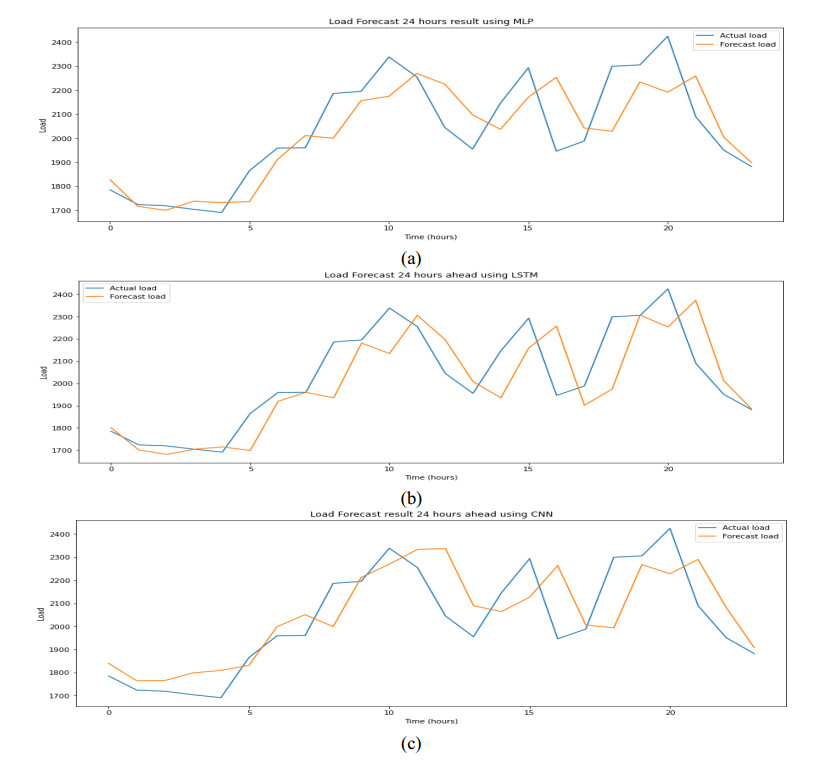
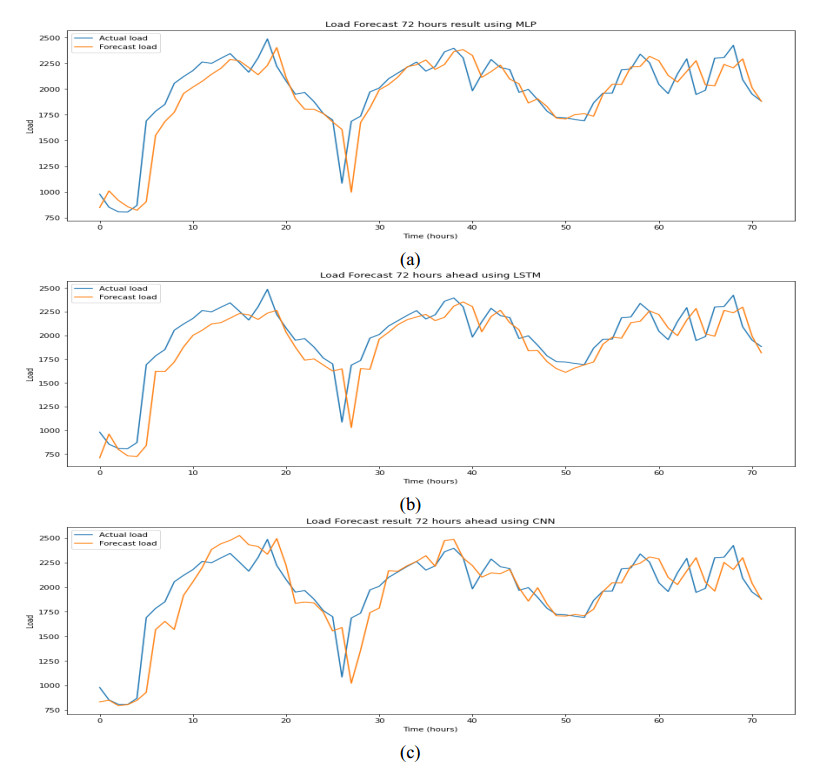
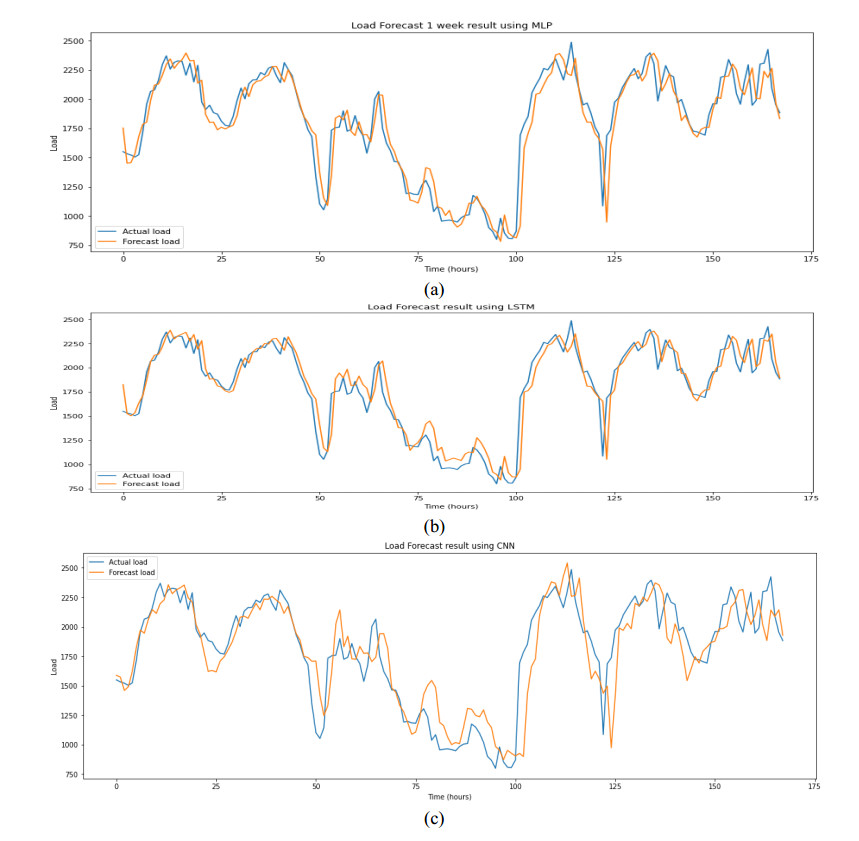
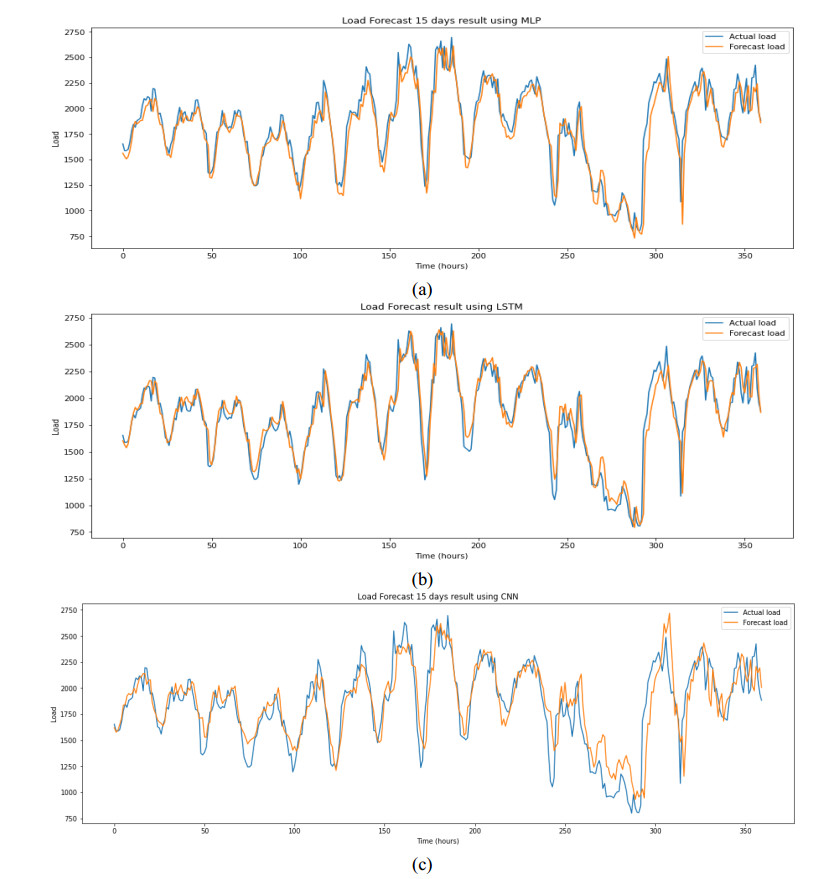
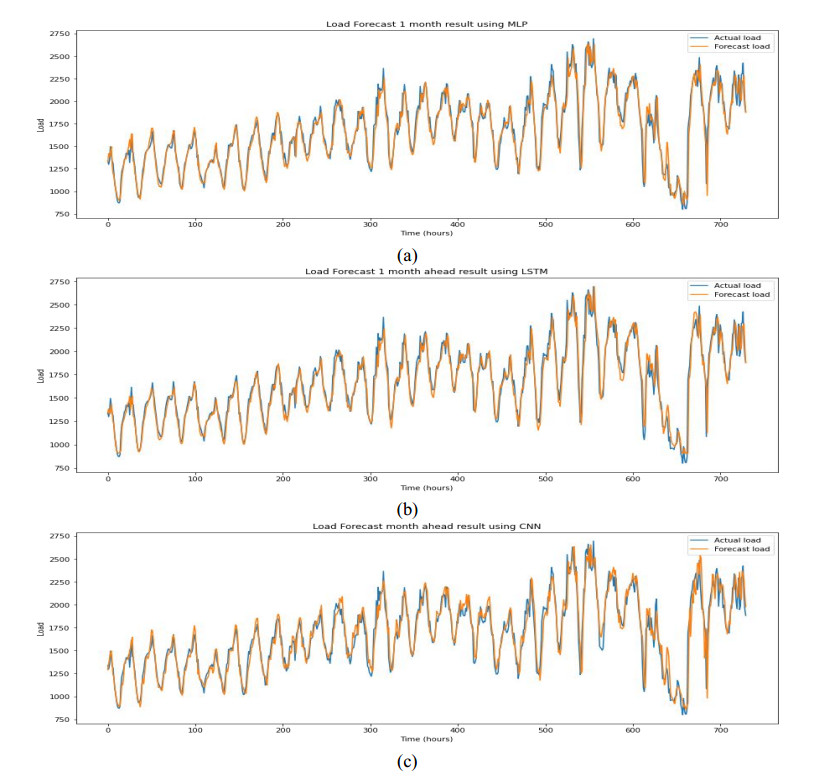
Figures(9) / Tables(6)

Faisal Mehmood Butt, Lal Hussain, Anzar Mahmood, Kashif Javed Lone. Artificial Intelligence based accurately load forecasting system to forecast short and medium-term load demands[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 400-425. doi: 10.3934/mbe.2021022







 DownLoad:
DownLoad: