Citation: Zhenlei Yang, Jie Wang, Ruixi Zhu. Identification of driver genes with aberrantly alternative splicing events in pediatric patients with retinoblastoma[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 328-338. doi: 10.3934/mbe.2021017

| [1] |
J. Reinhard, N. Wagner, M. M. Kramer, M. Jarocki, S. C. Joachim, H. B. Dick, et al., Expression Changes and Impact of the Extracellular Matrix on Etoposide Resistant Human Retinoblastoma Cell Lines, Int. J. Mol. Sci., 21 (2020), 4322. doi: 10.3390/ijms21124322

|
| [2] | I. Aerts, L. Lumbroso-Le Rouic, M. Gauthier-Villars, H. Brisse, F. Doz, [Retinoblastoma update], Arch Pediatr., 23 (2015), 112-116. |
| [3] |
F. Salviat, M. Gauthier-Villars, M. Carton, N. Cassoux, L. Lumbroso-Le Rouic, C. Dehainault, et al., Association Between Genotype and Phenotype in Consecutive Unrelated Individuals With Retinoblastoma, JAMA Ophthalmol., 138 (2020), 843-850. doi: 10.1001/jamaophthalmol.2020.2100
|
| [4] | P. R. Mendoza, H. E. Grossniklaus, The Biology of Retinoblastoma, in Progress in Molecular Biology and Translational Science, 2015,503-516. |
| [5] |
D. Lohmann, Retinoblastoma, Adv. Exp. Med. Biol., 685 (2010), 220-227. doi: 10.1007/978-1-4419-6448-9_21
|
| [6] |
M. Mehyar, M. Mosallam, A. Tbakhi, A. Saab, I. Sultan, R. Deebajah, et al., Impact of RB1 gene mutation type in retinoblastoma patients on clinical presentation and management outcome, Hematol. Oncol. Stem Cell Ther., 13 (2020), 152-159. doi: 10.1016/j.hemonc.2020.02.006
|
| [7] |
T. T. Kivela, T. Hadjistilianou, Neonatal Retinoblastoma, Asia. Pac. J. Oncol. Nurs., 4 (2017), 197-204. doi: 10.4103/apjon.apjon_18_17
|
| [8] |
P. Indovina, F. Pentimalli, N. Casini, I. Vocca, A. Giordano, RB1 dual role in proliferation and apoptosis: cell fate control and implications for cancer therapy, Oncotarget, 6 (2015), 17873-17890. doi: 10.18632/oncotarget.4286
|
| [9] |
L. Zheng, W. H. Lee, Retinoblastoma tumor suppressor and genome stability, Adv. Cancer Res., 85 (2002), 13-50. doi: 10.1016/S0065-230X(02)85002-3
|
| [10] |
D. E. Rushlow, B. M. Mol, J. Y. Kennett, S. Yee, S. Pajovic, B. L. Theriault, et al., Characterisation of retinoblastomas without RB1 mutations: genomic, gene expression, and clinical studies, Lancet Oncol., 14 (2013), 327-334. doi: 10.1016/S1470-2045(13)70045-7
|
| [11] |
C. Rodriguez-Martin, F. Cidre, A. Fernandez-Teijeiro, G. Gomez-Mariano, L. de la Vega, P. Ramos, et al., Familial retinoblastoma due to intronic LINE-1 insertion causes aberrant and noncanonical mRNA splicing of the RB1 gene, J. Hum.Genet., 61 (2016), 463-466. doi: 10.1038/jhg.2015.173
|
| [12] |
S. C. W. Lee, O. Abdel-Wahab, Therapeutic targeting of splicing in cancer, Nat. Med., 22 (2016), 976-986. doi: 10.1038/nm.4165
|
| [13] |
F. Supek, B. Minana, J. Valcarcel, T. Gabaldon, B. Lehner, Synonymous mutations frequently act as driver mutations in human cancers, Cell, 156 (2014), 1324-1335. doi: 10.1016/j.cell.2014.01.051
|
| [14] |
H. Jung, D. Lee, J. Lee, D. Park, Y. J. Kim, W. Y. Park, et al., Intron retention is a widespread mechanism of tumor-suppressor inactivation, Nat. Genet., 47 (2015), 1242-1248. doi: 10.1038/ng.3414
|
| [15] |
V. Spina, D. Rossi, Overview of non-coding mutations in chronic lymphocytic leukemia, Mol. Oncol., 13 (2019), 99-106. doi: 10.1002/1878-0261.12416
|
| [16] |
R. Karni, E. de Stanchina, S. W. Lowe, R. Sinha, D. Mu, A. R. Krainer, The gene encoding the splicing factor SF2/ASF is a proto-oncogene, Nat. Struct. Mol. Biol., 14 (2007), 185-193. doi: 10.1038/nsmb1209
|
| [17] |
M. Cohen-Eliav, R. Golan-Gerstl, Z. Siegfried, C. L. Andersen, K. Thorsen, T. F. Orntoft, et al., The splicing factor SRSF6 is amplified and is an oncoprotein in lung and colon cancers, J. Pathol., 229 (2013), 630-639. doi: 10.1002/path.4129
|
| [18] | R. Jia, C. Li, J. P. McCoy, C. X. Deng, Z. M. Zheng, SRp20 is a proto-oncogene critical for cell proliferation and tumor induction and maintenance, Int. J. Biol. Sci., 6 (2010), 806-826. |
| [19] | E. G. Bechara, E. Sebestyen, I. Bernardis, E. Eyras, J. Valcarcel, RBM5, 6, and 10 differentially regulate NUMB alternative splicing to control cancer cell proliferation, Mol. Cell, 52 (2013), 720-733. |
| [20] |
S. Rajasekaran, L. D. Nagarajha Selvan, K. Dotts, R. Kumar, P. Rishi, V. Khetan, et al., Non-coding and Coding Transcriptional Profiles Are Significantly Altered in Pediatric Retinoblastoma Tumors, Front. Oncol., 9 (2019), 221. doi: 10.3389/fonc.2019.00221
|
| [21] |
Y. Liao, G. K. Smyth, W. Shi, featureCounts: an efficient general purpose program for assigning sequence reads to genomic features, Bioinformatics, 30 (2014), 923-930. doi: 10.1093/bioinformatics/btt656
|
| [22] |
S. Shen, J. W. Park, Z. X. Lu, L. Lin, M. D. Henry, Y. N. Wu, et al., rMATS: robust and flexible detection of differential alternative splicing from replicate RNA-Seq data, Proc. Nat. Acad. Sci. U. S. A., 111 (2014), E5593-5601. doi: 10.1073/pnas.1419161111
|
| [23] |
M. I. Love, W. Huber, S. Anders, Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2, Genome Biol., 15 (2014), 550. doi: 10.1186/s13059-014-0550-8
|
| [24] |
C. Gu, X. Shi, Z. Huang, J. Chen, J. Yang, J. Shi, et al., A comprehensive study of construction and analysis of competitive endogenous RNA networks in lung adenocarcinoma, Biochim. Biophys. Acta Proteins Proteomics, 1868 (2020), 140444. doi: 10.1016/j.bbapap.2020.140444
|
| [25] |
X. Shi, T. Huang, J. Wang, Y. Liang, C. Gu, Y. Xu, et al., Next-generation sequencing identifies novel genes with rare variants in total anomalous pulmonary venous connection, EBioMedicine, 38 (2018), 217-227. doi: 10.1016/j.ebiom.2018.11.008
|
| [26] |
G. Yu, L. G. Wang, Y. Han, Q. Y. He, clusterProfiler: an R package for comparing biological themes among gene clusters, OMICS, 16 (2012), 284-287. doi: 10.1089/omi.2011.0118
|
| [27] |
V. Imperatore, A. M. Pinto, E. Gelli, E. Trevisson, V. Morbidoni, E. Frullanti, et al., Parent-of-origin effect of hypomorphic pathogenic variants and somatic mosaicism impact on phenotypic expression of retinoblastoma, Eur. J. Hum. Genet., 26 (2018), 1026-1037. doi: 10.1038/s41431-017-0054-6
|
| [28] |
C. J. Dommering, B. M. Mol, A. C. Moll, M. Burton, J. Cloos, J. C. Dorsman, et al., RB1 mutation spectrum in a comprehensive nationwide cohort of retinoblastoma patients, J. Med. Genet., 51 (2014), 366-374. doi: 10.1136/jmedgenet-2014-102264
|
| [29] |
J. Alonso, P. Garcia-Miguel, J. Abelairas, M. Mendiola, E. Sarret, M. T. Vendrell, et al., Spectrum of germline RB1 gene mutations in Spanish retinoblastoma patients: Phenotypic and molecular epidemiological implications, Hum. Mutat., 17 (2001), 412-422. doi: 10.1002/humu.1117
|
| [30] |
R. Condorelli, L. Spring, J. O'Shaughnessy, L. Lacroix, C. Bailleux, V. Scott, et al., Polyclonal RB1 mutations and acquired resistance to CDK 4/6 inhibitors in patients with metastatic breast cancer, Ann. Oncol., 29 (2018), 640-645. doi: 10.1093/annonc/mdx784
|
| [31] | F. Sanchez-Sanchez, M. Kruetzfeldt, C. Najera, S. Mittnacht, A novel constitutional mutation affecting splicing of retinoblastoma tumor suppressor gene intron 23 causes partial loss of pRB activity, Hum. Mutat., 25 (2005), 223. |
| [32] |
H. Dimaras, T. W. Corson, Retinoblastoma, the visible CNS tumor: A review, J. Neurosci. Res., 97 (2019), 29-44. doi: 10.1002/jnr.24213
|
| [33] |
H. Sun, Y. Wang, M. Chinnam, X. Zhang, S. W. Hayward, B. A. Foster, et al., E2f binding-deficient Rb1 protein suppresses prostate tumor progression in vivo, Proc. Nat. Acad. Sci. U. S. A., 108 (2011), 704-709. doi: 10.1073/pnas.1015027108
|
| [34] | F. P. M. Cremers, W. Lee, R. W. J. Collin, R. Allikmets, Clinical spectrum, genetic complexity and therapeutic approaches for retinal disease caused by ABCA4 mutations, Prog. Retinal Eye Res., 2020, 100861, forthcoming. |
| [35] |
C. P. Pang, D. S. Lam, Differential occurrence of mutations causative of eye diseases in the Chinese population, Hum. Mutat., 19 (2002), 189-208. doi: 10.1002/humu.10053
|
| [36] |
T. Chen, W. Zheng, J. Chen, S. Lin, Z. Zou, X. Li, Z. Tan, Systematic analysis of survival-associated alternative splicing signatures in clear cell renal cell carcinoma, J. Cell. Biochem., 121 (2020), 4074-4084. doi: 10.1002/jcb.29590
|
| [37] |
M. Yu, W. Hong, S. Ruan, R. Guan, L. Tu, B. Huang, et al., Genome-Wide Profiling of Prognostic Alternative Splicing Pattern in Pancreatic Cancer, Front. Oncol., 9 (2019), 773. doi: 10.3389/fonc.2019.00773
|
| [38] |
V. Prima, S. P. Hunger, Cooperative transformation by MEF2D/DAZAP1 and DAZAP1/MEF2D fusion proteins generated by the variant t(1;19) in acute lymphoblastic leukemia, Leukemia, 21 (2007), 2470-2475. doi: 10.1038/sj.leu.2404962
|
| [39] |
R. Choudhury, S. G. Roy, Y. S. Tsai, A. Tripathy, L. M. Graves, Z. Wang, The splicing activator DAZAP1 integrates splicing control into MEK/Erk-regulated cell proliferation and migration, Nat. Commun., 5 (2014), 3078. doi: 10.1038/ncomms4078
|
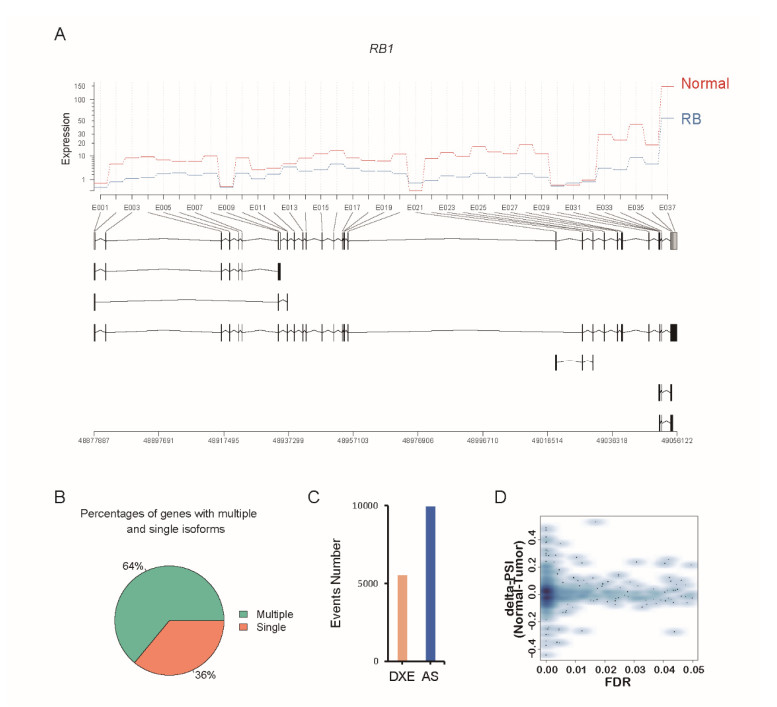




Figures(5)

Zhenlei Yang, Jie Wang, Ruixi Zhu. Identification of driver genes with aberrantly alternative splicing events in pediatric patients with retinoblastoma[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 328-338. doi: 10.3934/mbe.2021017







 DownLoad:
DownLoad: