Citation: Arturo J. Nic-May, Eric J. Avila-Vales. Global dynamics of a two-strain flu model with a single vaccination and general incidence rate[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7862-7891. doi: 10.3934/mbe.2020400

| [1] | World Health Organization. Influenza (Seasonal), 6 November 2018. Available from: https://www.who.int/es/news-room/fact-sheets/detail/influenza-(seasonal). |
| [2] |
A. Rahman, X. Zou, Flu epidemics: a two strain flu model with a single vaccination, J. Biol. Dyn., 5 (2011), 376-390. doi: 10.1080/17513758.2010.510213

|
| [3] |
M. Medina, E. Vintiñi, J. Villena, R. Raya, S. Alvarez, Lactococcus lactisas an adjuvant and delivery vehicle of antigens against pneumococcal respiratory infections, Bioeng. Bugs, 1 (2010), 313-325. doi: 10.4161/bbug.1.5.12086
|
| [4] |
G. Chowell, C. Ammon, N. Hengartner, J. Hyman, Transmission dynamics of the great influenza pandemic of 1918 in Geneva, Switzerland: Assessing the effects of hypothetical interventions, J. Theor. Biol., 241 (2006), 193-204. doi: 10.1016/j.jtbi.2005.11.026
|
| [5] |
C. Mills, J. Robins, M. Lipsitch, Transmissibility of 1918 pandemic influenza, Nature, 432 (2004), 904-906. doi: 10.1038/nature03063
|
| [6] |
S. Cauchemez, A. Valleron, P. Boëlle, A. Flahault, N. Ferguson, Estimating the impact of school closure on influenza transmission from Sentinel data, Nature, 452 (2008), 750-754. doi: 10.1038/nature06732
|
| [7] |
V. Capasso, G. Serio, A Generalization of the Kermack-Mckendrick deterministic epidemic model, Math. Biosci., 42 (1978), 43-61. doi: 10.1016/0025-5564(78)90006-8
|
| [8] |
I. Baba, E. Hincal, A model for influenza with vaccination and awareness, Chaos, Solitons, Fractals, 106 (2018), 49-55. doi: 10.1016/j.chaos.2017.11.003
|
| [9] |
I. Baba, E. Hincal, Global stability analysis of two-strain epidemic model with bilinear and nonmonotone incidence rates, Eur. Phys. J. Plus, 132 (2017), 208. doi: 10.1140/epjp/i2017-11476-x
|
| [10] |
A. Korobeinikov, P. K. Maini, Non-linear incidence and stability of infectious disease models, Math. Med. Biol., 22 (2005), 113-128. doi: 10.1093/imammb/dqi001
|
| [11] | L. Wang, X. Zhang, Z. Liu, An SEIR Epidemic Model with Relapse and General Nonlinear Incidence Rate with Application to Media Impact, Qual. Theory Dyn. Syst., 17 (2017), 309-329. |
| [12] |
P. Van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29-48. doi: 10.1016/S0025-5564(02)00108-6
|
| [13] |
H. R. Thieme, Persistence under relaxed point-dissipativity (with application to an endemic model), SIAM J. Math. Anal., 24 (1993), 407-435. doi: 10.1137/0524026
|
| [14] | G. J. Butler, H. I. Freedman, P. Waltman, Uniformly persistent systems, Proc. Am. Math. Soc., 96 (1989), 425-430. |
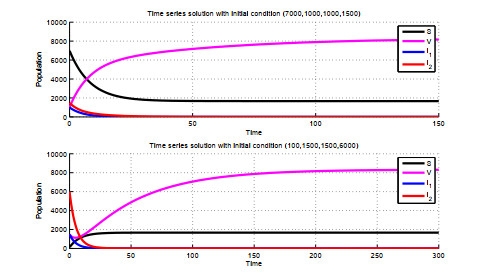
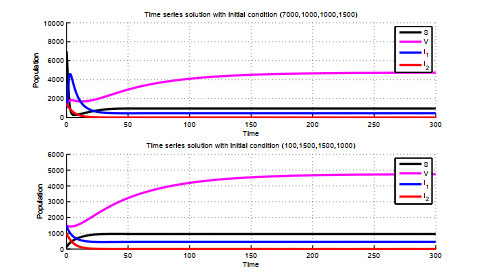
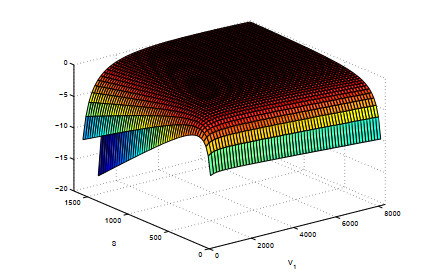
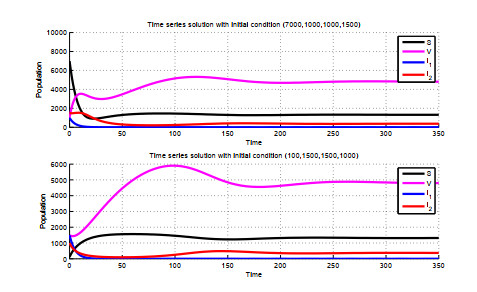

Figures(5)

Arturo J. Nic-May, Eric J. Avila-Vales. Global dynamics of a two-strain flu model with a single vaccination and general incidence rate[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7862-7891. doi: 10.3934/mbe.2020400







 DownLoad:
DownLoad: