Citation: Yueping Dong, Jianlu Ren, Qihua Huang. Dynamics of a toxin-mediated aquatic population model with delayed toxic responses[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5907-5924. doi: 10.3934/mbe.2020315

| [1] | C. H. Walker, R. Sibly, S. Hopkin, D. B. Peakall, Principles of Ecotoxicology, CRC Press, Boca Raton, 2012. |
| [2] | The Canadian Council of Ministers of the Environment, Canadian water quality guidelines for the protection of aquatic life: Guidance on the site-specific application of water quality guidelines in Canada: procedures for deriving numerical water quality objectives, 2003, http://ceqgrcqe.ccme.ca/download/en/221. |
| [3] | US national archives and records administration, code of federal regulations, title 40-protection of environment, Appendix A to part 423-126 priority pollutants. |
| [4] | S. M. Bartell, R. A. Pastorok, H. R. Akcakaya, H. Regan, S. Ferson, C. Mackay, Realism and relevance of ecological models used in chemical risk assessment, Hum. Ecol. Risk Assess., 9 (2003), 907-938. |
| [5] | R. A. Pastorok, S. M. Bartell, S. Ferson, L. R. Ginzburg, Ecological Modeling in Risk Assessment: Chemical Effects on Populations, Ecosystems, and Landscapes, CRC Press, Boca Raton, 2001. |
| [6] | T. G. Hallam, C. E. Clark, R. R. Lassiter, Effects of toxicants on populations: Aqualitative approach I. Equilibrium environmental exposure, Ecol. Model., 18 (1983), 291-304. |
| [7] | T. G. Hallam, C. E. Clark, G. S. Jordan, Effects of toxicants on populations: A qualitative approach II. First order kinetics, J. Math. Biol., 18 (1983), 25-37. |
| [8] | T. G. Hallam, J. T. De Luna, Effects of toxicants on populations: A qualitative approach III. Environmental and food chain pathways, J. Theor. Biol., 109 (1984), 411-429. |
| [9] | J. T. De Luna, T. G. Hallam, Effects of toxicants on populations: A qualitative approach IV. Resource-consumer-toxicant models, Ecol. Model., 109 (1987), 249-273. |
| [10] | Q. Huang, L. Parshotam, H. Wang, C. Bampfylde, M. A. Lewis, A model for the impact of contaminaants on fish population dynamics, J. Theor. Biol., 334 (2013), 71-79. |
| [11] | Q. Huang, G. Seo, C. Shan, Bifurcations and global dynamics in a toxin-dependent aquatic population model, Math. Biosci., 296 (2018), 26-35. |
| [12] | N. MacDonald, Biological Delay Systems: Linear Stability Theory, Cambridge University Press, Cambridge, 1989. |
| [13] | Y. Kuang, Delay Differential Equations: With Applications in Population Dynamics, Academic Press, 1993. |
| [14] | J. M. Cushing, Integro-differential Equations and Delay Models in Population Dynamics, Springer Science & Business Media, 2013. |
| [15] | S. Ruan, Absolute stability, conditional stability and bifurcation in Kolmogorov-type predatorprey systems with discrete delays, Quart. Appl. Math, 59 (2001), 159-173. |
| [16] | J. Chattopadhayay, R. R. Sarkar, S. Mandal, Toxin-producing plankton may act as a biological control for planktonic blooms-field study and mathematical modelling, J. Theor. Biol., 215 (2002), 333-344. |
| [17] | J. Chattopadhayay, R. R. Sarkar, A. El Abdllaoui, A delay differential equation model on harmful algal blooms in the presence of toxic substances, IMA J. Math. Appl. Med. Biol., 19 (2002), 137-161. |
| [18] | Y. Dong, G. Huang, R. Miyazaki, Y. Takeuchi, Dynamics in a tumor immune system with time delays, Appl. Math. Comput., 252 (2015), 99-113. |
| [19] | Z. Jiang, L. Wang, Global Hopf bifurcation for a predator-prey system with three delays, Int. J. Bifur. Chaos, 27 (2017), 1750108. |
| [20] | Y. Dong, Y. Takeuchi, S. Nakaoka, A mathematical model of multiple delayed feedback control system of the gut microbiota-Antibiotics injection controlled by measured metagenomic data, Nonlinear Anal. Real World Appl., 43 (2018), 1-17. |
| [21] | Y. Dong, M. Sen, M. Banerjee, Y. Takeuchi, S. Nakaoka, Delayed feedback induced complex dynamics in an Escherichia coli and Tetrahymena system, Nonlinear Dyn., 94 (2018), 1447-1466. |
| [22] | R. K. Upadhyay, S. Mishra, Y. Dong, Y. Takeuchi, Exploring the dynamics of a tritrophic food chain model with multiple gestation periods, Math. Biosci. Eng., 16 (2019), 4660-4691. |
| [23] | Z. Jiang, L. Wang, Global Hopf bifurcation of a delayed phytoplankton-zooplankton system considering toxin producing effect and delay dependent coeffcient, Math. Biosci. Eng., 16 (2019), 3807-3829. |
| [24] | P. Panja, S. K. Mondal, D. K. Jana, Effects of toxicants on Phytoplankton-Zooplankton-Fish dynamics and harvesting, Chaos Soliton. Fract., 104 (2017), 389-399. |
| [25] | S. K. Sahani, How delay can affect the survival of species in polluted environment, in "Soft Computing for Problem Solving" (eds. J. C. Bansal, K. N. Das, A. Nagar, K. Deep, A. K. Ojha), Springer, Singapore, (2019), 913-923. |
| [26] | V. Vasconcelos, J. Azevedo, M. Silva, V. Ramos, Effects of marine toxins on the reproduction and early stages development of aquatic organisms, Mar. Drugs, 8 (2010), 59-79. |
| [27] | L. Casarini, A. Franchini, D. Malagoli, E. Ottaviani, Evaluation of the effects of the marine toxin okadaic acid by using FETAX assay, Toxicol. Lett., 169 (2007), 145-151. |
| [28] | M. A. Beketov, M. Liess, Acute and delayed effects of the neonicotinoid insecticide thiacloprid on seven freshwater arthropods, Environ. Toxicol. Chem., 27 (2008), 461-470. |
| [29] | A. C. Gutleb, J. Appelman, M. C. Bronkhorst, J. H. J. van den Berg, A. Spenkelink, A. Brouwer et al., Delayed effects of pre- and early-life time exposure to polychlorinated biphenyls on tadpoles of two amphibian species (Xenopus lae6is and Rana temporaria), Environ. Toxicol. Pharmacol., 8 (1999), 1-14. |
| [30] | Q. Huang, H. Wang, M. A. Lewis, The impact of environmental toxins on predator-prey dynamics, J. Theor. Biol., 378 (2015), 12-30. |
| [31] | C. Shan, Q. Huang, Direct and indirect effects of toxins on competition dynamics of species in an aquatic environment, J. Math. Biol., 78 (2019), 739-766. |
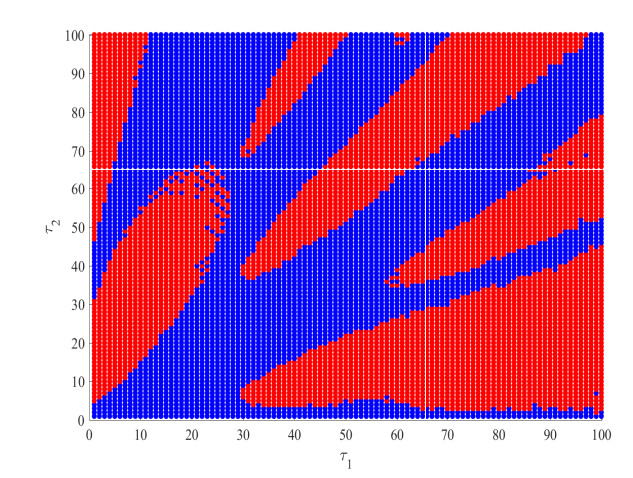
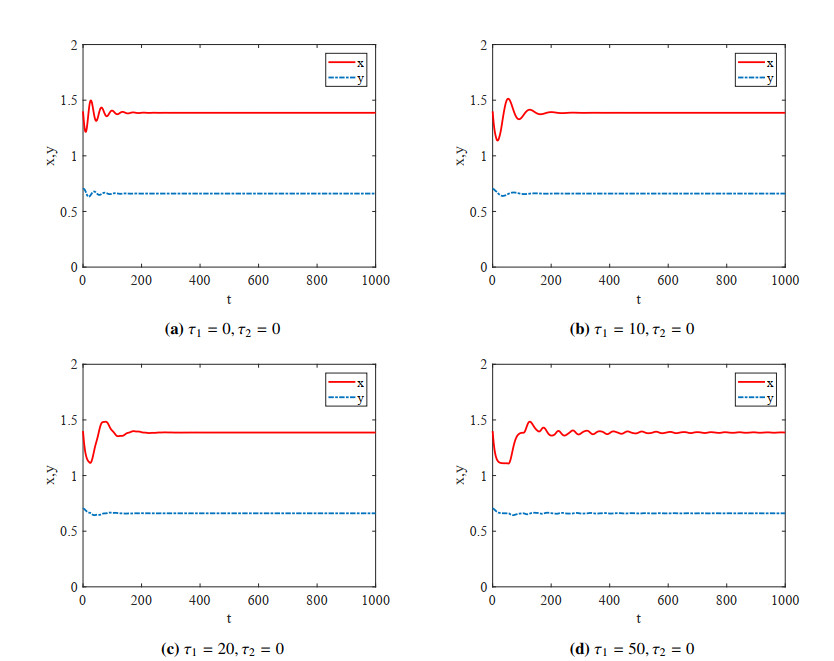
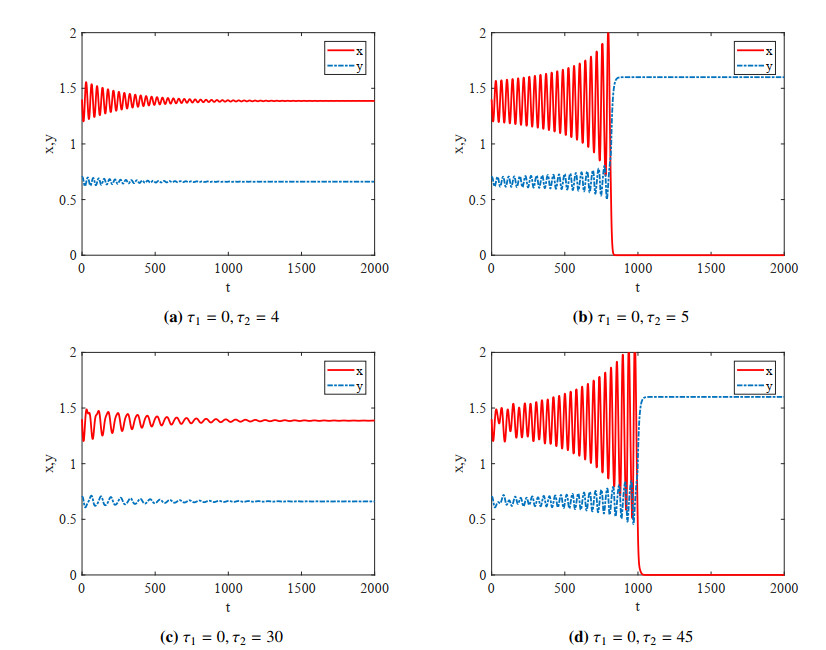
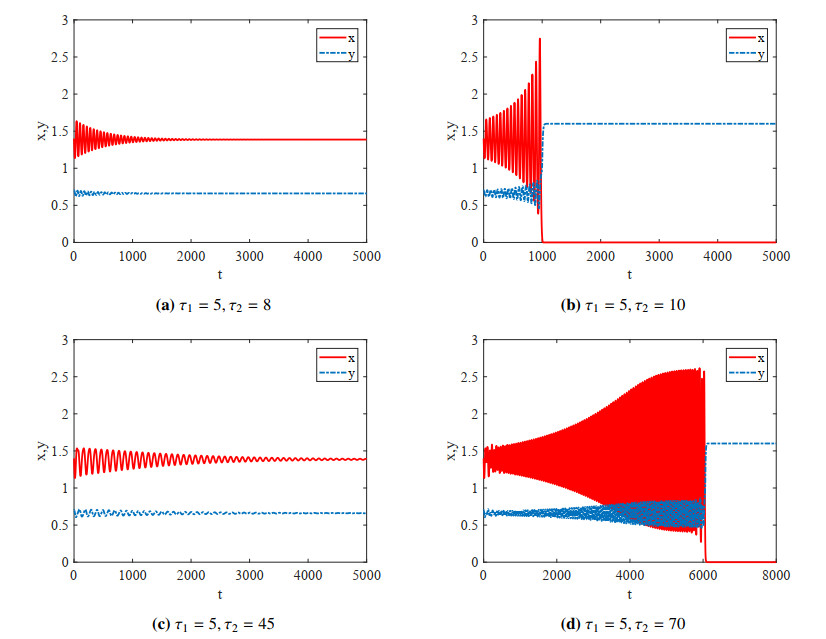
Figures(4)

Yueping Dong, Jianlu Ren, Qihua Huang. Dynamics of a toxin-mediated aquatic population model with delayed toxic responses[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5907-5924. doi: 10.3934/mbe.2020315







 DownLoad:
DownLoad: