Citation: Dongmei Li, Tana Guo, Yajing Xu. The effects of impulsive toxicant input on a single-species population in a small polluted environment[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 8179-8194. doi: 10.3934/mbe.2019413

| [1] | Z. Jiang, Can we prevent extinction?, Bull. Sci. Technol., 61 (2016), 1978–1982. |
| [2] | Q. Zhao, G. Huang and Y. Ma, The ecological environment conditions and construction of an ecological civilization in China, Acta Ecol. Sin., 36 (2016), 6328–6335. |
| [3] | T. G. Hallam, C. E. Clark and R. R. Lassider, Effects of toxicants on populations: A qualitative approach I equilibrium environmental exposure, Eool. Model, 8 (1983), 291–304. |
| [4] | T. G. Hallam, C. E. Clark and G. S. Jordan, Effects of toxicants on populations: A qualitative approach Ⅱ first order kinetics, Math. Biol., 18 (1983), 25–37. |
| [5] | T. G. Hallam and J. T. Deluna, Effects of toxicants on populations: A qualitative approachc. environmental and food chain pathways, J. Theor. Biol., 109 (1984), 411–429. |
| [6] | F. Wei, A. H. Geritz and J. Cai, A stochastic single-species population model with partial pollution tolerance in a polluted environment, Appl. Math. Lett., 63 (2017), 130–136. |
| [7] | Y. Feng and K. Wang, The survival analysis for single-species system in a polluted environment, J. Ann. Diff. Equ., 22 (2006), 154–159. |
| [8] | J. He and K. Wang, The survival analysis for a single-species population model in a polluted environment, Nonlinear Anal.: Real World Appl., 10 (2009), 1555–1571. |
| [9] | J. He and K. Wang, The survival analysis for a population in a polluted environment, Appl. Math. Model., 31 (2007), 2227–2238. |
| [10] | Z. Luo, Y. Yu and Z. Ba, The thresholds of survival for a single-species population model in a polluted environment, J. Biomath., 26 (2011), 211–222. |
| [11] | A. Kumar, A. K. Agrawal, A. Hasan, et al., Modeling the effect of toxicant on the deformity in a subclass of a biological species, Model. Earth Syst. Environ., 2 (2016), 40–40. |
| [12] | V. Ducrot, E. Billoir, J. Garric, et al., From individual to population level effects of toxicants in the tubicifid branchiura sowerbyi using threshold effect models in a bayesian framework, Environ. Sci. Technol., 44 (2010), 3566–3571. |
| [13] | B. Liu, L. Chen and Y. Zhang, The effects of impulsive toxicant input on a population in a polluted environment, J. Biol. Syst., 11 (2003), 265–274. |
| [14] | H. Zhang, X. Meng and L. Chen, Impact of impulsive effect on a population dynamical behaviorin a polluted environment, J. Dalian Univ. Technol., 48 (2008), 147–153. |
| [15] | L. Bao and X. Ban, The study on single population model with pulse input of environmental toxin, J. Jiangxi Norm. Univ. (Nat. Sci.), 36 (2012), 502–505. |
| [16] | B. Liu, L. Xu and B. Kang, Dynamics of a stage structured pest control model in a polluted environment with pulse pollution input, J. Appl. Math., 2013 (2009), 1–12. |
| [17] | J. Jiao and L. Chen, The extinction threshold on a single population model with pulse input of environmental toxin in a polluted environment, Math. Appl., 22 (2009), 11–19. |
| [18] | Y. Xue, J Wang and Z. Jin, The persistent threshold of single population under pulse input of environment, Wseas Trans. Math., 1 (2007), 22–29. |
| [19] | Y. Hou and J. Jiao, Dynamical analysis of a single population model with delayed growth response and pulse input in polluted environment, Math. Pract. Theory, 47 (2017), 119–127. |
| [20] | X. Song, H. Guo and X. Shi, The Theory of Impulsive Differential Equation and Its Application, Science Press, Beijing, 2011. |
| [21] | Y. Xiao, Y. Zhou and S. Tang, Biological Mathematical Principles, Xi'an Jiaotong University Press, Xi'an, 2012. |
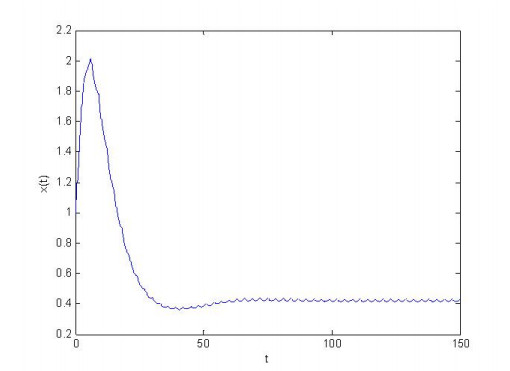
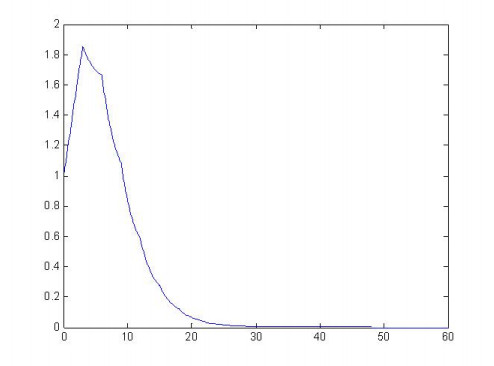
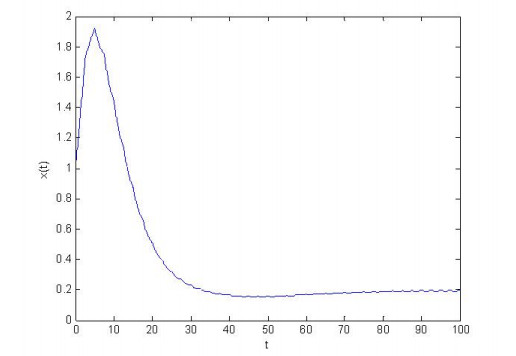
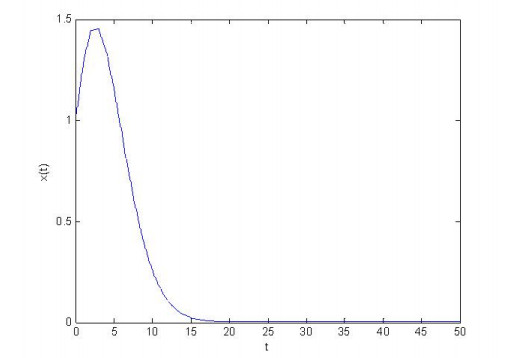
Figures(4) / Tables(1)

Dongmei Li, Tana Guo, Yajing Xu. The effects of impulsive toxicant input on a single-species population in a small polluted environment[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 8179-8194. doi: 10.3934/mbe.2019413







 DownLoad:
DownLoad: