Citation: Yendry N. Arguedas, Mario Santana-Cibrian, Jorge X. Velasco-Hernández. Transmission dynamics of acute respiratory diseases in a population structured by age[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7477-7493. doi: 10.3934/mbe.2019375

| [1] | C. Ruiz-Matus, P. Kuri-Morales and J.Narro-Robles, Comportamiento de las temporadas de in-fluenza en México de 2010 a 2016, análisis y prospectiva, Gaceta Médica de México, 153 (2017), 205–213. |
| [2] | B. Cowling, G. Freeman, J. Wong, et al., Preliminary inferences on the age-specific seriousness of human disease caused by avian influenza a (H7N9) infections in China, March to April 2013, Eur. Surveil., 18 (2013), 20475. |
| [3] | G. Emukule, P. Spreeuwenberg, S. Chaves, et al., Estimating influenza and respiratory syncytial virus-associated mortality in Western Kenya using health and demographic surveillance system data, 2007-2013, PloS One, 12 (2017), e0180890. |
| [4] | S. Tempia, S. Walazaa, C. Viboud, et al., Mortality associated with seasonal and pandemic in-fluenza and respiratory syncytial virus among children < 5 years of age in a high HIV prevalence setting in South Africa, 1998–2009, Clin. Infect. Dis., 58 (2014), 1241–1249. |
| [5] | L. White, M. Waris, P. Cane, et al., The transmission dynamics of groups a and b human respi-ratory syncytial virus (hRSV) in England & Wales and Finland: seasonality and cross-protection, Epidemiol. Infect., 133 (2005), 279–289. |
| [6] | H. Zhou, W. Thompson, C. Viboud, et al., Hospitalizations associated with influenza and respira-tory syncytial virus in the United States, 1993–2008, Clin. Infect. Dis., 54 (2012), 1427–1436. |
| [7] | A. Comas-García, C. García-Sepúlveda, J. Méndez-de Lira, et al., Mortality attributable to pan-demic influenza a (H1N1) 2009 in San Luis Potosí, México, Influenza Other Respir. Viruses, 5 (2011), 76–82. |
| [8] | CJ. Worby, C. Kenyon, R. Lynfield, et al., Examining the role of different age groups, and of vaccination during the 2012 Minnesota pertussis outbreak, Sci. Rep., 5 (2015), 13182. |
| [9] | C. Nj and K. Subbarao, Influenza, Lancet, 354 (1999), 1277–82. |
| [10] | S. Towers and Z. Feng, Social contact patterns and control strategies for influenza in the elderly, Math. Biosci., 240 (2012), 241–249. |
| [11] | S. Cauchemez, A. Valleron, P. Boelle, et al., Estimating the impact of school closure on influenza transmission from sentinel data, Nature, 452 (2008), 750. |
| [12] | J. Tamerius, C. Viboud, J. Shaman, et al., Impact of school cycles and environmental forcing on the timing of pandemic influenza activity in Mexican states, May-December 2009, PLoS Comput. Biol., 11 (2015), e1004337. |
| [13] | M. Angulo and J. Velasco-Hernandez, Robust qualitative estimation of time-varying contact rates in uncertain epidemics, Epidemics (2018). Available from: https://doi.org/10.1016/j. epidem.2018.03.001 |
| [14] | A. Elkadry, Transmission rate in partial differential equation in epidemic models, Ph.D thesis, Marshall University, 2013. |
| [15] | K. Hadeler, Parameter identification in epidemic models, Math. Biosci., 229 (2011), 185–189. |
| [16] | A. Lange, Reconstruction of disease transmission rates: applications to measles, dengue, and influenza, J. Theor. Biol., 400 (2016), 138–153. |
| [17] | M. Pollicott, H. Wang and H. Weiss, Extracting the time-dependent transmission rate from infec-tion data via solution of an inverse ODE problem, J. Biol. Dyn., 6 (2012), 509–523. |
| [18] | R. Anderson and R. May, Infectious diseases of humans: dynamics and control, Oxford University Press, 1992. |
| [19] | J. Velasco-Hernández, M. Nuñez-Lopez, A. Comas-García, et al., Superinfection between in-fluenza and RSV alternating patterns in San Luis Potosí State, México, PloS One, 10 (2015), e0115674. |
| [20] | R. Schmidt-Ott, M. Schwehm and M. Eichner, Influence of social contact patterns and demo-graphic factors on influenza simulation results, BMC Infect. Dis., 16 (2016), 646. |
| [21] | A. Agresti, Categorical data analysis, 2nd edition, John Wiley & Sons, 2002. |
| [22] | J. Ponciano and M. Capistrán, First principles modeling of nonlinear incidence rates in sea-sonalepidemics, PLoS Comput. Biol., 7 (2011), e1001079. |
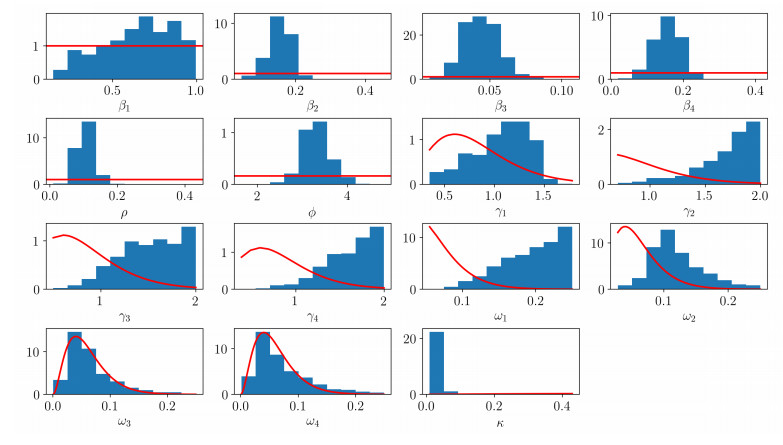
| [23] | J. Christen and C. Fox., A general purpose sampling algorithm for continuous distributions (the t-walk), Bayesian Anal., 5 (2010), 263–282. |
| [24] | C. Robert and G. Casella, Monte Carlo Statistical Methods, 2nd edition, Springer, New York, 2004. |
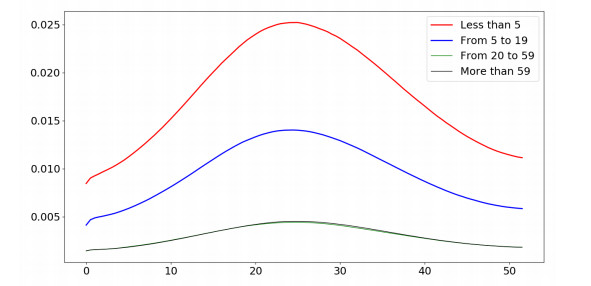
| [25] | J. Mossong, N. Hens, M. Jit, et al., Social contacts and mixing patterns relevant to the spread of infectious diseases, PLoS Med., 5 (2008), e74. |
Figures(6) / Tables(7)

Yendry N. Arguedas, Mario Santana-Cibrian, Jorge X. Velasco-Hernández. Transmission dynamics of acute respiratory diseases in a population structured by age[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7477-7493. doi: 10.3934/mbe.2019375







 DownLoad:
DownLoad: