Citation: Sourav Kumar Sasmal, Jeet Banerjee, Yasuhiro Takeuchi. Dynamics and spatio-temporal patterns in a prey–predator system with aposematic prey[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 3864-3884. doi: 10.3934/mbe.2019191

| [1] | R. M. May, Biological populations with nonoverlapping generations: stable points, stable cycles, and chaos, Science, 186 (1974), 645–647. |
| [2] | J. Banerjee, S. K. Sasmal and R. K. Layek, Supercritical and subcritical Hopf-bifurcations in a two-delayed prey–predator system with density-dependent mortality of predator and strong Allee effect in prey, BioSystems, 180 (2019), 19–37. |
| [3] | S. K. Sasmal, Population dynamics with multiple Allee effects induced by fear factors–A mathe-matical study on prey–predator interactions, Appl. Math. Model., 64 (2018), 1–14. |
| [4] | J. Banerjee, T. Ranjan and R. K. Layek, Dynamics of cancer progression and suppression: A novel evolutionary game theory based approach, 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), (2015), 5367–5371. |
| [5] | J. Banerjee, T. Ranjan and R. K. Layek, Stability Analysis of Population Dynamics Model in |
| [6] | Microbial Biofilms with Non-participating Strains, 7th ACM International Conference on Bioin-formatics, Computational Biology, and Health Informatics, (201, 220–230. |
| [7] | 6. M. Wang, A. L. Schaefer, A. A. Dandekar, et al., Quorum sensing and policing of Pseudomonas aeruginosa social cheaters, Proceed. Nat. Aca. Sci., 112 (2015), 2182191. |
| [8] | 7. J. Banerjee, R. K. Layek, S. K. Sasmal, et al., Delayed evolutionary model for public goods competition with policing in phenotypically-variant bacterial biofilms, Europhys. Let., (2019) (in press). |
| [9] | 8. M. Stevens and G. D. Ruxton, Linking the evolution and form of warning coloration in nature, Proceed. Royal Soc. B Biol. Sci., 27(2011), 417–426. |
| [10] | 9. J. Skelhorn, C. G. Halpin and C. Rowe, Learning about aposematic prey, Behav. Ecol., 27 (2016), 955–964. |
| [11] | 10. J. Skelhorn and C. Rowe, Predators' toxin burdens influence their strategic decisions to eat toxic prey, Curr. Biol., 17 (2007), 1479–1483. |
| [12] | 11. C. G. Halpin, J. Skelhorn and C. Rowe, Predators' decisions to eat defended prey depend on the size of undefended prey, Animal Behav., 85 (2013), 1315–1321. |
| [13] | 12. K. E. Smith, C. G. Halpin and C. Rowe, Body size matters for aposematic prey during predator aversion learning, Behav. Process., 109 (2014), 173–179. |
| [14] | 13. J. E. Huheey, Mathematical models of mimicry, Am. Nat., 131 (1988), S22–S41. |
| [15] | 14. J. C. Santos, L. A. Coloma and D. C. Cannatella, Multiple, recurring origins of aposematism and diet specialization in poison frogs, Proceed. Nat. Aca. Sci., 100 (2003), 12792–12797. |
| [16] | 15. R. A. Saporito, R. Zuercher, M. Roberts, et al., Experimental evidence for aposematism in the dendrobatid poison frog Oophaga pumilio, Copeia, 2007 (2007), 1006–1011. |
| [17] | 16. J. C. Santos and D. C. Cannatella, Phenotypic integration emerges from aposematism and scale in poison frogs, Proceed. Nat. Aca. Sci., 108 (2011), 6-6180. |
| [18] | 17. E. D. Brodie III, Differential avoidance of coral snake banded patterns by free-ranging avian predators in Costa Rica, Evolution, 47 (1993), 227–235. |
| [19] | 18. Y. Takeuchi, W. Wang, S. Nakaoka, et al., Dynamical adaptation of parental care, Bull. Math. Biol., 71 (2009), 931–951. |
| [20] | 19. S. Chakraborty, P. K. Tiwari, S. K. Sasmal, et al., Interactive effects of prey refuge and additional food for predator in a diffusive predator-prey system, Appl. Math. Model., 47 (7), 128–140. |
| [21] | 20. D. Alonso, F. Bartumeus and J. Catalan, Mutual interference between predators can give rise to Turing spatial patterns, Ecology, 83 (2002), 28–34. |
| [22] | 21. J. R. Meyer, S. P. Ellner, N. G. Hairston, et al., Prey evolution on the time scale of predator–prey dynamics revealed by allele-specific quantitative PCR, Proceed. Nat. Aca. Sci., 103 (2006), 10690–10695. |
| [23] | 22. L. Lindström, R. V. Alatalo, J. Mappes, et al., Can aposematic signals evolve by gradual change?, Nature, 397 (1999), 249–251. |
| [24] | 23. J. Gohli and G. Högstedt, Explaining the evolution of warning coloration: Secreted secondary defence chemicals may facilitate the evolution of visual aposematic signals, PLoS One, 4 (2009), e5779. |
| [25] | 24. J. B. Barnett, C. Michalis, N. E. Scott-Samuel, et al., Distance-dependent defensive coloration in the poison frog Dendrobates tinctorius, Dendrobatidae, Proceed. Nat. Aca. Sci., 115 (2018), 6416–6421. |
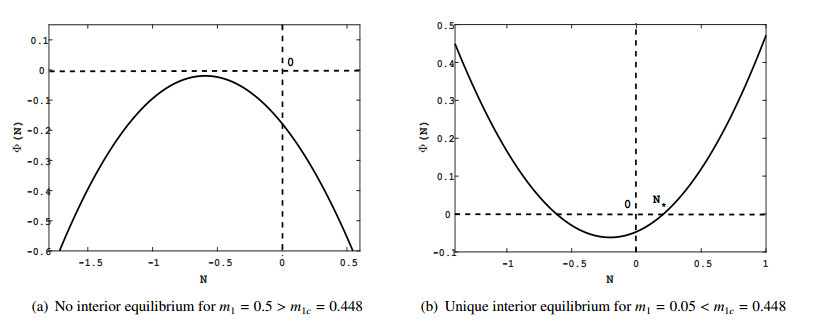
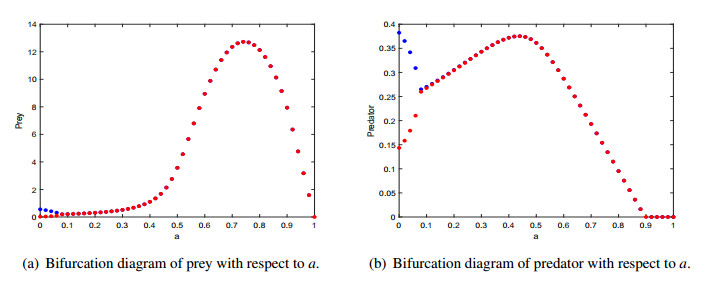
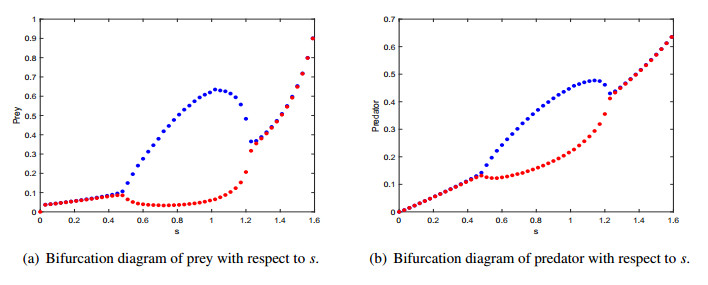
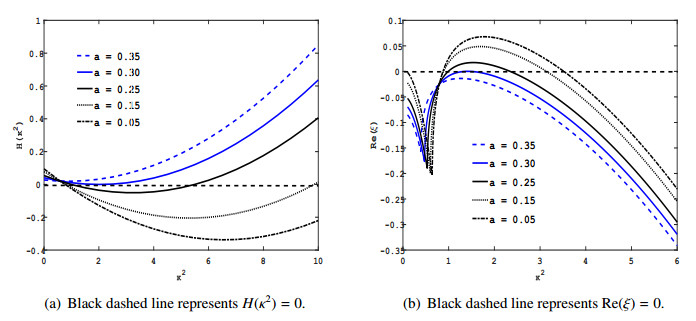

Figures(9) / Tables(2)

Sourav Kumar Sasmal, Jeet Banerjee, Yasuhiro Takeuchi. Dynamics and spatio-temporal patterns in a prey–predator system with aposematic prey[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 3864-3884. doi: 10.3934/mbe.2019191







 DownLoad:
DownLoad: