Citation: Md Masud Rana, Chandani Dissanayake, Lourdes Juan, Kevin R. Long, Angela Peace. Mechanistically derived spatially heterogeneous producer-grazer model subject to stoichiometric constraints[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 222-233. doi: 10.3934/mbe.2019012

| [1] | T. Andersen (1997) Pelagic nutrient cycles; herbivores as sources and sinks. Springer, Berlin. |
| [2] | T. Andersen , J. J. Elser and D. O. Hessen, Stoichiometry and population dynamics. Ecol. Lett., 7 (2004), 884–900. |
| [3] | S. A. Berger, S. Diehl, T. J. Kunz, D. Albrecht, A. M. Oucible and S. Ritzer, Light supply, plankton biomass, and seston stoichiometry in a gradient of lake mixing depths. Limnol. Oceanogr., 51 (2006), 1898–1905. |
| [4] | E. L. Cussler (1996) Diffusion: Mass Transfer in Fluid Systems. Cambridge University Press. |
| [5] | S. Diehl, S. Berger and R. W¨ohrl, Flexible nutrient stoichiometry mediates environmental influences on phytoplankton and its resources. Ecology , 86 (2005), 2931–2945. |
| [6] | C. Dissanayake (2016) Finite element simulation of space/time behavior in a two species ecological stoichiometric model. PhD thesis, Texas Tech University. |
| [7] | C. Dissanayake, L. Juan, K. R. Long, A. Peace and M. M. Rana (under review 2018) Genotypic selection in spatially heterogeneous producer-grazer systems subject to stoichiometric constraints. Bulletin of Mathematical Biology. |
| [8] | J. Huisman, J. Sharples, J. M. Stroom, P. M. Visser, W. E. A. Kardinaal, J. M. Verspagen and Sommeijer B, Changes in turbulent mixing shift competition for light between phytoplankton species. Ecology, 85 (2004), 2960–2970. |
| [9] | J. T. Kirk (1994) Light and photosynthesis in aquatic ecosystems. Cambridge University Press. |
| [10] | D. Kuefler, T. Avgar and J. M. Fryxell, Rotifer population spread in relation to food, density and predation risk in an experimental system. J. Anim. Ecol., 81 (2012), 323–329. |
| [11] | D. Kuefler, T. Avgar and J. M. Fryxell, Density-and resource-dependent movement characteristics in a rotifer. Funct. Ecol., 27 (2013), 323–328. |
| [12] | I. Loladze, Y. Kuang and J. J. Elser, Stoichiometry in producer–grazer systems: linking energy flow with element cycling. Bull. Math. Biol., 62 (2000), 1137–1162. |
| [13] | A. Lorke, Investigation of turbulent mixing in shallow lakes using temperature microstructure measurements. Aquat. Sci.-Research Across Boundaries, 60 (1998), 210–219. |
| [14] | A. Peace, H.Wang and Y. Kuang, Dynamics of a producer–grazer model incorporating the effects of excess food nutrient content on grazer's growth. Bull. Math. Biol., 76 (2014), 2175–2197. |
| [15] | F. H. Shu (1991) The Physics of Astrophysics, Vol. 2: Radiation. Univ. Sci. Books, Mill Valley CA. |
| [16] | R. W. Sterner and J. J. Elser (2002) Ecological stoichiometry: the biology of elements from molecules to the biosphere. Princeton University Press. |
| [17] | H. Wang, H. L. Smith, Y. Kuang and J. J. Elser, Dynamics of stoichiometric bacteria-algae interactions in the epilimnion. SIAM J. Appl. Math., 68 (2007), 503–522. |
| [18] | H. Wang, Y. Kuang and I. Loladze, Dynamics of a mechanistically derived stoichiometric producer-grazer model. J. Biol. Dynam., 2 (2008), 286–296. |
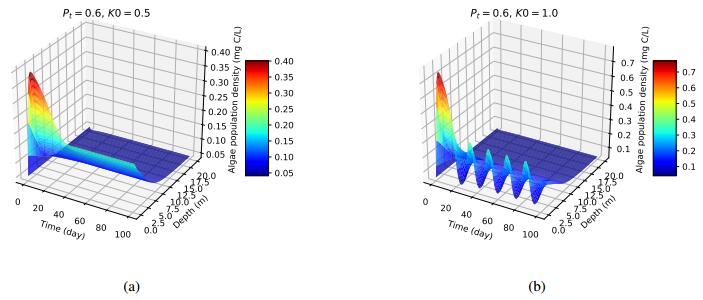
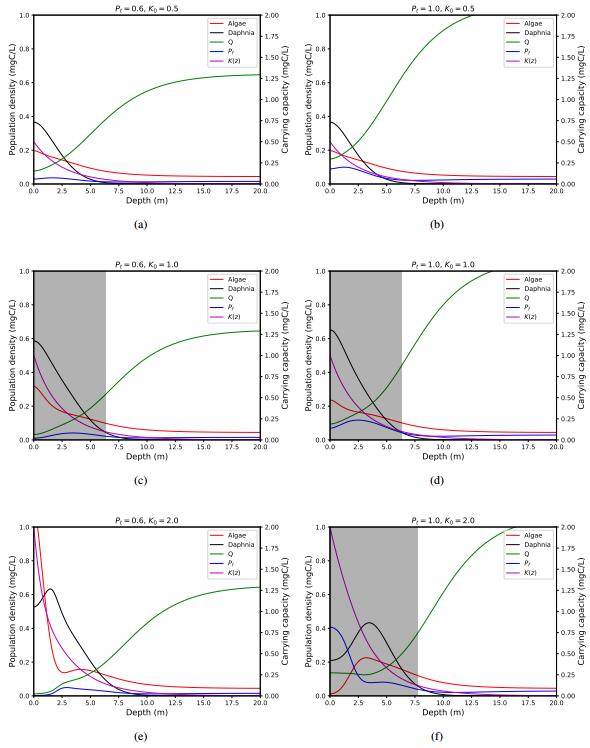
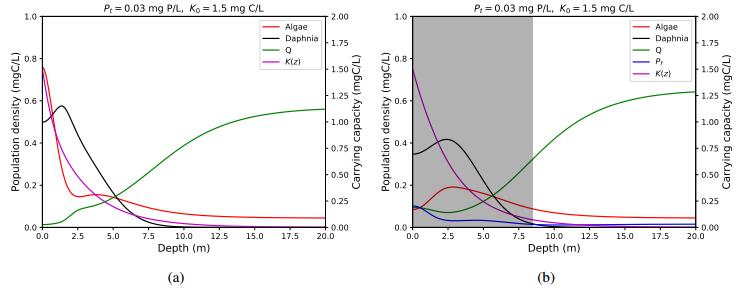
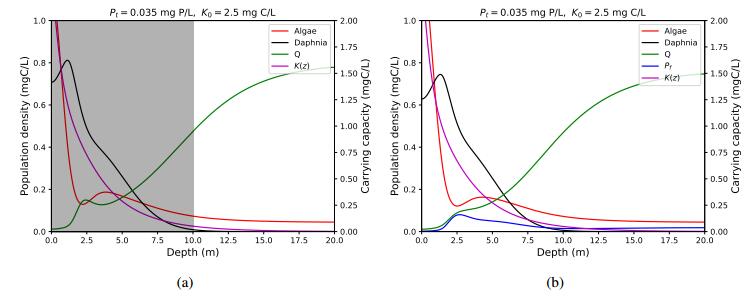
Figures(4) / Tables(1)

Md Masud Rana, Chandani Dissanayake, Lourdes Juan, Kevin R. Long, Angela Peace. Mechanistically derived spatially heterogeneous producer-grazer model subject to stoichiometric constraints[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 222-233. doi: 10.3934/mbe.2019012







 DownLoad:
DownLoad: