Citation: Bo Zheng, Wenliang Guo, Linchao Hu, Mugen Huang, Jianshe Yu. Complex wolbachia infection dynamics in mosquitoes with imperfect maternal transmission[J]. Mathematical Biosciences and Engineering, 2018, 15(2): 523-541. doi: 10.3934/mbe.2018024

| [1] | [ G. Bian,Y. Xu,P. Lu,Z. Xi, The endosymbiotic bacterium Wolbachia induces resistance to dengue virus in Aedes aegypti, PLoS Pathogens, 6 (2010): e1000833. |
| [2] | [ G. Bian,D. Joshi,Y. Dong,P. Lu,G. Zhou,X. Pan,Y. Xu,G. Dimopoulos,Z. Xi, Wolbachia invades Anopheles stephensi populations and induces refractoriness to plasmodium infection, Science, 340 (2013): 748-751. |
| [3] | [ L. B. Carrington,J. R. Lipkowitz,A. A. Hoffmann,M. Turelli, A re-examination of Wolbachia-induced cytoplasmic incompatibility in California Drosophila simulans, PLoS One, 6 (2011): e22565. |
| [4] | [ E. Caspari,G. S. Watson, On the evolutionary importance of cytoplasmic sterility in mosquitoes, Evolution, 13 (1959): 568-570. |
| [5] | [ H. L. Dutra,L. M Dos Santos,E. P. Caragata,J. B. Silva,D. A. Villela,R. Maciel-De-Freitas, From lab to field: The influence of urban landscapes on the invasive potential of Wolbachia in Brazilian Aedes aegypti mosquitoes, PLoS Neglected Tropical Diseases, 9 (2015): e0003689. |
| [6] | [ H. L. Dutra,M. N. Rocha,F. B. Dias,S. B. Mansur,E. P. Caragata,L. A. Moreira, Wolbachia, blocks currently circulating Zika virus isolates in Brazilian Aedes aegypti, Cell Host Microbe, 19 (2016): 771-774. |
| [7] | [ S. L. Dobson,C. W. Fox,F. M. Jiggins, The effect of Wolbachia-induced cytoplasmic incompatibility on host population size in natural and manipulated systems, Proceedings of the Royal Society B Biological Sciences, 269 (2002): 437-445. |
| [8] | [ J. Z. Farkas,P. Hinow, Structured and unstructured continuous models for Wolbachia infections, Bulletin of Mathematical Biology, 72 (2010): 2067-2088. |
| [9] | [ P. E. Fine, On the dynamics of symbiote-dependent cytoplasmic incompatibility in culicine mosquitoes, Journal of Invertebrate Pathology, 31 (1978): 10-18. |
| [10] | [ F. D. Frentiu,T. Zakir,T. Walker,J. Popovici,A. T. Pyke,D. H. A. Van, Limited dengue virus replication in field-collected Aedes aegypti mosquitoes infected with Wolbachia, PLoS Neglected Tropical Diseases, 8 (2014): e2688. |
| [11] | [ C. A. Hamm,D. J. Begun,A. Vo,C. C. Smith,P. Saelao,A. O. Shaver,J. Jaenike,M. Turelli, Wolbachia do not live by reproductive manipulation alone: Infection polymorphism in Drosophila suzukii and D. subpulchrella, Molecular Ecology, 23 (2014): 4871-4885. |
| [12] | [ R. Haygood,M. Turelli, Evolution of incompatibility-inducing microbes in subdivided host populations, Evolution, 63 (2009): 432-447. |
| [13] | [ M. W. Hirsch, S. Smale and R. L. Devaney, Differential Equations, Dynamical Systems, And An Introduction to Chaos Second edition. Pure and Applied Mathematics (Amsterdam), 60. Elsevier/Academic Press, Amsterdam, 2004. |
| [14] | [ A. A. Hoffmann,M. Turelli, Unidirectional incompatibility in Drosophila simulans: Inheritance, geographic variation and fitness effects, Genetics, 119 (1988): 435-444. |
| [15] | [ A. A. Hoffmann, M. Turelli and L. G. Harshman, Factors affecting the distribution of cytoplasmic incompatibility in Drosophila simulans, Genetics, 126 (1991), 933–948. |
| [16] | [ A. A. Hoffmann and M. Turelli, Cytoplasmic incompatibility in insects, In Influential Passengers: Inherited Microorganisms and Arthropod Reproduction, S. L. O'Neill et al. , eds, Oxford University Press, Oxford, (1997), 42–80. |
| [17] | [ A. A. Hoffmann,B. L. Montgomery,J. Popovici,I. Iturbeormaetxe,P. H. Johnson,F. Muzzi,M. Greenfield,M. Durkan,Y. S. Leong,Y. Dong,H. Cook,J. Axford, J. Axford, Gallahan, N. Kenny, C. Omodei, E. A. McGraw, P. A. Ryan, S. A. Ritchie, M. Turelli and S. L. O'Neill, Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission, Nature, 476 (2011): 454-459. |
| [18] | [ A. A. Hoffmann,I. Iturbeormaetxe,A. G. Callahan,B. L. Phillips,K. Billington,J. K. Axford,B. Montgomery,A. P. Turley,S. L. O'Neill, Stability of the wMel Wolbachia infection following Invasion into Aedes aegypti populations, PLoS Neglected Tropical Diseases, 8 (2014): e3115-e3115. |
| [19] | [ L. Hu,M. Huang,M. Tang,J. Yu,B. Zheng, Wolbachia spread dynamics in stochastic environments, Theoretical Population Biology, 106 (2015): 32-44. |
| [20] | [ M. Huang,M. Tang,Y. Yu, Wolbachia infection dynamics by reaction-diffusion equations, Science China Mathematics, 58 (2015): 77-96. |
| [21] | [ M. Huang,J. Yu,L. Hu,B. Zheng, Qualitative analysis for a Wolbachia infection model with diffusion, Science China Mathematics, 59 (2016): 1249-1266. |
| [22] | [ M. J. Keeling,F. M. Jiggins,J. M. Read, The invasion and coexistence of competing Wolbachia strains, Heredity, 91 (2003): 382-388. |
| [23] | [ P. Kriesner,A. A. Hoffmann,S. F. Lee,M. Turelli,A. R. Weeks, Rapid sequential spread of two Wolbachia variants in Drosophila simulan, PLoS Pathogens, 9 (2013): e1003607. |
| [24] | [ P. Kriesner,W. R. Conner,A. R. Weeks,M. Turelli,A. A. Hoffmann, Wolbachia infection frequency cline in Drosophila melanogaster and the possible role of reproductive dormancy, Evolution, 70 (2016): 979-997. |
| [25] | [ C. J. Mcmeniman,R. V. Lane,B. N. Cass,A. W. Fong,M. Sidhu,Y. F. Wang,S. L. O'Neill, Stable introduction of a life-shortening Wolbachia infection into the mosquito Aedes aegypti, Science, 323 (2009): 141-144. |
| [26] | [ T. H. Nguyen,H. L. Nguyen,T. Y. Nguyen,S. N. Vu,N. D. Tran,T. N. Le,S. Kutcher,T. P. Hurst,T. T. Duong,J. A. Jeffery,J. M. Darbro,B. H. Kay,I. Iturbe-Ormaetxe,J. Popovici,B. L. Montgomery,A. P. Turley,F. Zigterman,H. Cook,P. E. Cook,P. H. Johnson,P. A. Ryan,C. J. Paton,S. A. Ritchie,C. P. Simmons,L. O'Neill,A. A. Hoffmann, Field evaluation of the establishment potential of wMelPop Wolbachia in Australia and Vietnam for dengue control, Parasites Vectors, 8 (2015): p563. |
| [27] | [ J. L. Rasgon,L. M. Styer,T. W. Scott, Wolbachia-induced mortality as a mechanism to modulate pathogen transmission by vector arthropods, Journal of Medical Entomology, 40 (2003): 125-132. |
| [28] | [ P. A. Ross,I. Wiwatanaratanabutr,J. K. Axford,V. L. White,N. M. Endersby-Harshman,A. A. Hoffmann, Wolbachia infections in Aedes aegypti differ markedly in their response to cyclical heat stress, PLoS Pathogens, 13 (2017): e1006006. |
| [29] | [ J. G. Schraiber,A. N. Kaczmarczyk,R. Kwok,M. Park,R. Silverstein,F. U. Rutaganira,T. Aggarwal,M. A. Schwemmer,C. L. Hom,R. K. Grosberg,S. J. Schreiber, Constraints on the use of lifespan-shortening Wolbachia to control dengue fever, Journal of Theoretical Biology, 297 (2012): 26-32. |
| [30] | [ M. Turelli,A. A. Hoffmann, Rapid spread of an inherited incompatibility factor in California Drosophila, Nature, 353 (1991): 440-442. |
| [31] | [ M. Turelli, Evolution of incompatibility-inducing microbes and their hosts, Evolution, 48 (1994): 1500-1513. |
| [32] | [ M. Turelli,A. A. Hoffmann, Cytoplasmic incompatibility in Drosophila simulans: Dynamics and parameter estimates from natural populations, Genetics, 140 (1995): 1319-1338. |
| [33] | [ M. Turelli, Microbe-induced cytoplasmic incompatibility as a mechanism for introducing transgenes into arthropod populations, Insect Molecular Biology, 8 (1999): 243-255. |
| [34] | [ M. Turelli, Cytoplasmic incompatibility in populations with overlapping generations, Evolution, 64 (2010): 232-241. |
| [35] | [ T. Walker,P. H. Johnson,L. A. Moreira,I. Iturbeormaetxe,F. D. Frentiu,C. J. McMeniman,Y. S. Leong,Y. Dong,J. Axford,P. Kriesner,A. L. Lloyd,S. A. Ritchie,S. L. O'Neill,A. A. Hoffmann, The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations, Nature, 476 (2011): 450-453. |
| [36] | [ A. R. Weeks,M. Turelli,W. R. Harcombe,K. T. Reynolds,A. A. Hoffmann, From parasite to mutualist: Rapid evolution of Wolbachia in natural populations of Drosophila, PLoS Biology, 5 (2007): e114-e114. |
| [37] | [ Z. Xi,S. L. Dobson, Characterization of Wolbachia transfection efficiency by using microinjection of embryonic cytoplasm and embryo homogenate, Applied Environmental Microbiology, 71 (2005): 3199-3204. |
| [38] | [ Z. Xi,J. L. Dean,C. C. Khoo,S. L. Dobson, Generation of a novel Wolbachia infection in Aedes albopictus (Asian tiger mosquito) via embryonic microinjection, Insect Biochemistry Molecular Biology, 35 (2005): 903-910. |
| [39] | [ Z. Xi,C. C. Khoo,S. L. Dobson, Wolbachia establishment and invasion in an Aedes aegypti laboratory population, Science, 310 (2005): 326-328. |
| [40] | [ Z. Xi,C. C. Khoo,S. L. Dobson, Interspecific transfer of Wolbachia into the mosquito disease vector Aedes albopictus, Proceedings of the Royal Society B Biological Sciences, 273 (2006): 1317-1322. |
| [41] | [ B. Zheng,M. Tang,J. Yu, Modeling Wolbachia spread in mosquitoes through delay differential equations, SIAM Journal on Applied Mathematics, 74 (2014): 743-770. |
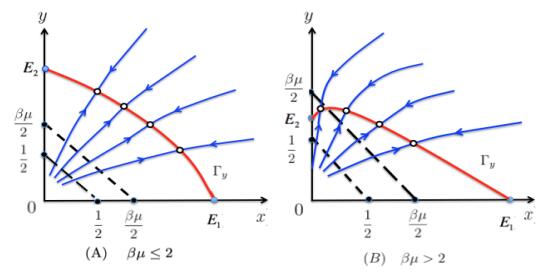
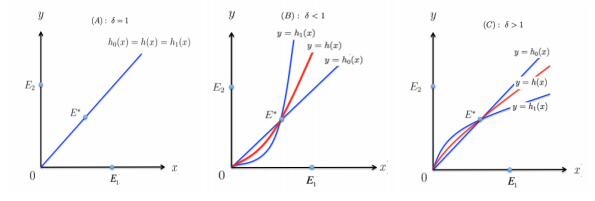
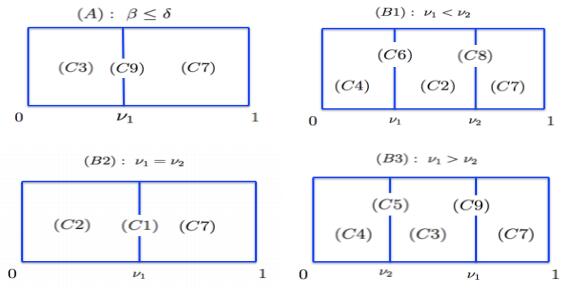
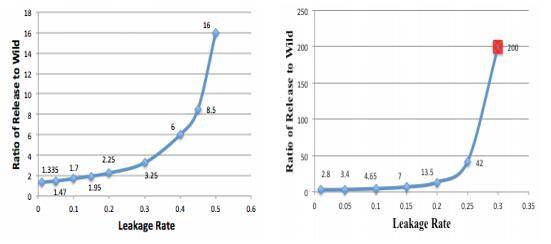
Figures(4)

Bo Zheng, Wenliang Guo, Linchao Hu, Mugen Huang, Jianshe Yu. Complex wolbachia infection dynamics in mosquitoes with imperfect maternal transmission[J]. Mathematical Biosciences and Engineering, 2018, 15(2): 523-541. doi: 10.3934/mbe.2018024







 DownLoad:
DownLoad: