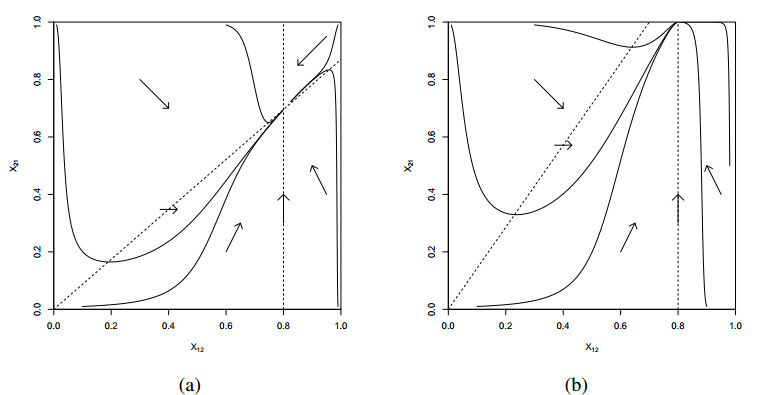
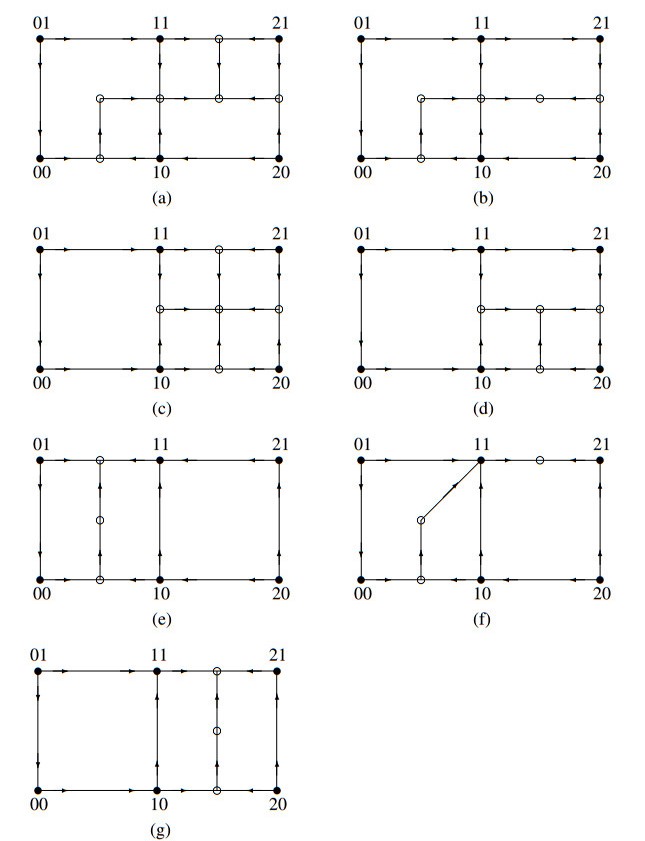
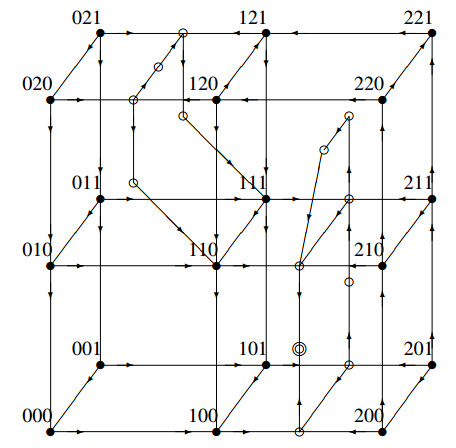
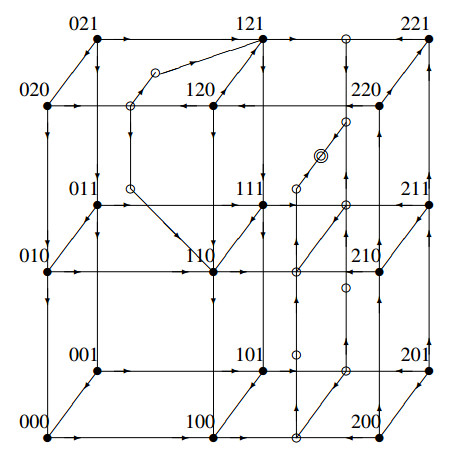
The Precursor Shutoff Valve (PSV) has been proposed as a motif in biochemical networks, active for example in prioritization of primary over secondary metabolism in plants in low-input conditions. Another branch prioritization mechanism in a biochemical network is a difference in thresholds for activation of the two pathways from the branch point. It has been shown by Adams and colleagues that both mechanisms can play a part in a model of plant metabolism involving Michaelis-Menten kinetics [
Citation: Roderick Edwards, Michelle Wood. Branch prioritization motifs in biochemical networks with sharp activation[J]. AIMS Mathematics, 2022, 7(1): 1115-1146. doi: 10.3934/math.2022066
The Precursor Shutoff Valve (PSV) has been proposed as a motif in biochemical networks, active for example in prioritization of primary over secondary metabolism in plants in low-input conditions. Another branch prioritization mechanism in a biochemical network is a difference in thresholds for activation of the two pathways from the branch point. It has been shown by Adams and colleagues that both mechanisms can play a part in a model of plant metabolism involving Michaelis-Menten kinetics [

| [1] |
Z. P. Adams, J. Ehlting, R. Edwards, The regulatory role of shikimate in plant phenylalanine metabolism, J. Theor. Biol., 462 (2019), 158–170. doi: 10.1016/j.jtbi.2018.11.005. doi: 10.1016/j.jtbi.2018.11.005

|
| [2] |
S. Collombet, C. van Oevelen, J. L. Sardina, W. Abou-Jaoudé, B. Di Stefano, M. Thomas-Chollier, et al., Logical modeling of lymphoid and myeloid cell specification and transdifferentiation, Proceedings of the National Academy of Sciences, 114 (2017), 5792–5799. doi: 10.1073/pnas.1610622114. doi: 10.1073/pnas.1610622114
|
| [3] |
M. I. Davidich, S. Bornholdt, Boolean network model predicts cell cycle sequence of fission yeast, PLoS ONE, 3 (2008), e1672. doi: 10.1371/journal.pone.0001672. doi: 10.1371/journal.pone.0001672
|
| [4] |
H. de Jong, J. Geiselmann, G. Batt, C. Hernandez, M. Page, Qualitative simulation of the initiation of sporulation in Bacillus subtilis, B. Math. Biol., 66 (2004), 261–299. doi: 10.1016/j.bulm.2003.08.009. doi: 10.1016/j.bulm.2003.08.009
|
| [5] |
H. de Jong, J. Geiselmann, C. Hernandez, M. Page, Genetic Network Analyzer: Qualitative simulation of genetic regulatory networks, Bioinformatics, 19 (2003), 336–344. doi: 10.1093/bioinformatics/btf851. doi: 10.1093/bioinformatics/btf851
|
| [6] |
S. Drulhe, G. Ferrari-Trecate, H. de Jong, The switching threshold reconstruction problem for piecewise-affine models of genetic regulatory networks, IEEE T. Automat. Contr., 53 (2008), 153–165. doi: 10.1109/TAC.2007.911326. doi: 10.1109/TAC.2007.911326
|
| [7] |
R. Edwards, Analysis of continuous-time switching networks, Physica D, 146 (2000), 165–199. doi: 10.1016/S0167-2789(00)00130-5. doi: 10.1016/S0167-2789(00)00130-5
|
| [8] |
R. Edwards, S. Kim, P. van den Driessche, Control design for sustained oscillation in a two gene regulatory network, J. Math. Biol., 62 (2011), 453–478. doi: 10.1007/s00285-010-0343-y. doi: 10.1007/s00285-010-0343-y
|
| [9] |
R. Edwards, A. Machina, G. McGregor, P. van den Driessche, A modelling framework for gene regulatory networks including transcription and translation, B. Math. Biol., 77 (2015), 953–983. doi: 10.1007/s11538-015-0073-9. doi: 10.1007/s11538-015-0073-9
|
| [10] |
L. Glass, R. Edwards, Hybrid models of genetic networks: Mathematical challenges and biological relevance, J. Theor. Biol., 458 (2018), 111–118. doi: 10.1016/j.jtbi.2018.09.014. doi: 10.1016/j.jtbi.2018.09.014
|
| [11] |
L. Glass, S.A. Kauffman, Co-operative components, spatial localization and oscillatory cellular dynamics, J. Theor. Biol., 34 (1972), 219–237. doi: 10.1016/0022-5193(72)90157-9. doi: 10.1016/0022-5193(72)90157-9
|
| [12] |
L. Glass, S. A. Kauffman, The logical analysis of continuous, non-linear biochemical control networks, J. Theor. Biol., 39 (1973), 103–129. doi: 10.1016/0022-5193(73)90208-7. doi: 10.1016/0022-5193(73)90208-7
|
| [13] |
J. L. Gouzé, T. Sari, A class of piecewise linear differential equations arising in biological models, Dynamical Systems, 17 (2002), 299–316. doi: 10.1080/1468936021000041681. doi: 10.1080/1468936021000041681
|
| [14] | F. Grognard, H. de Jong, J. L. Gouzé, Piecewise-linear models of genetic regulatory networks: Theory and example, in Biology and Control Theory: Current Challenges (eds. I. Queinnec, S. Tarbouriech, G. Garcia and S.I. Niculescu), Springer, 357 (2007), 137–159. doi: 10.1007/978-3-540-71988-5_7. |
| [15] |
L. Ironi, L. Panzeri, A computational framework for qualitative simulation of nonlinear dynamical models of gene-regulatory networks, BMC Bioinformatics, 10 (2009), S14. doi: 10.1186/1471-2105-10-S12-S14. doi: 10.1186/1471-2105-10-S12-S14
|
| [16] |
L. Ironi, D. X. Tran, Nonlinear and temporal multiscale dynamics of gene regulatory networks: A qualitative simulator, Math. Comput. Simul., 125 (2016), 15–37. doi: 10.1016/j.matcom.2015.11.007. doi: 10.1016/j.matcom.2015.11.007
|
| [17] |
L. Mendoza, D. Thieffry, E. R. Alvarez-Buylla, Genetic control of flower morphogenesis in Arabidopsis thaliana: a logical analysis, Bioinformatics, 15 (1999), 593–606. doi: 10.1093/bioinformatics/15.7.593. doi: 10.1093/bioinformatics/15.7.593
|
| [18] |
T. Mestl, E. Plahte, S. W. Omholt, A mathematical framework for describing and analysing gene regulatory networks, J. Theor. Biol., 176 (1995), 291–300. doi: 10.1006/jtbi.1995.0199. doi: 10.1006/jtbi.1995.0199
|
| [19] |
E. Plahte, S. Kjøglum, Analysis and generic properties of gene regulatory networks with graded response functions, Physica D, 201 (2005), 150–176. doi: 10.1016/j.physd.2004.11.014. doi: 10.1016/j.physd.2004.11.014
|
| [20] |
E. Plahte, T. Mestl, S.W. Omholt, A methodological basis for description and analysis of systems with complex switch-like interactions, J. Math. Biol., 36 (1998), 321–348. doi: 10.1007/s002850050103. doi: 10.1007/s002850050103
|
| [21] |
D. Ropers, H. de Jong, M. Page, D. Schneider, J. Geiselmann, Qualitative simulation of the carbon starvation response in Escherichia coli, Biosystems, 84 (2006), 124–152. doi: 10.1016/j.biosystems.2005.10.005. doi: 10.1016/j.biosystems.2005.10.005
|
Figures(11) / Tables(10)

Roderick Edwards, Michelle Wood. Branch prioritization motifs in biochemical networks with sharp activation[J]. AIMS Mathematics, 2022, 7(1): 1115-1146. doi: 10.3934/math.2022066







 DownLoad:
DownLoad: