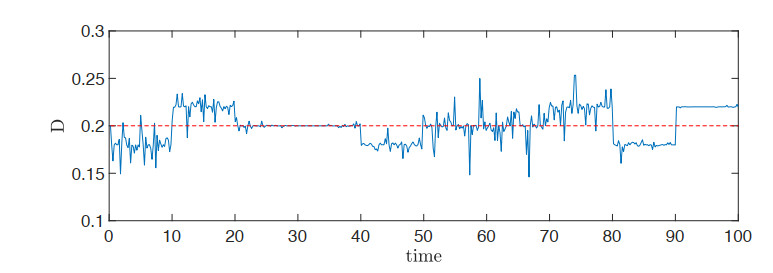
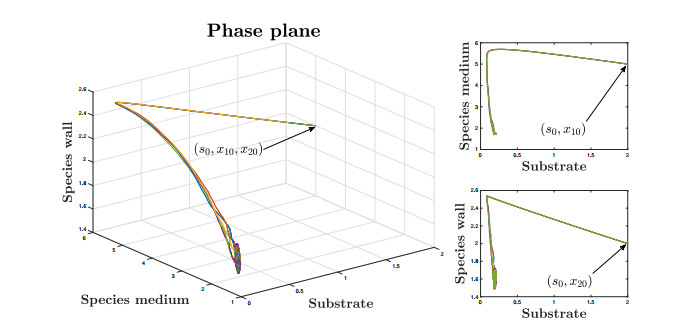
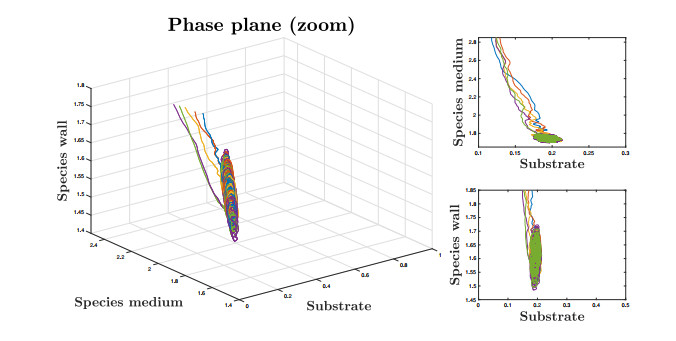
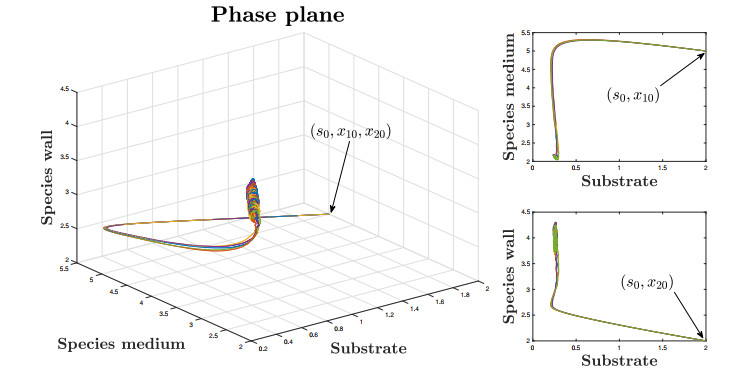
This paper investigates a chemostat model with wall growth and Haldane consumption kinetics. In addition, bounded random fluctuations on the input flow, which are modeled by means of the well-known Ornstein-Uhlenbeck process, are considered to obtain a much more realistic model fitting in a better way the phenomena observed by practitioners in real devices. Once the existence and uniqueness of global positive solution has been proved, as well as the existence of deterministic absorbing and attracting sets, the random dynamics inside the attracting set is studied in detail to provide conditions under which persistence of species is ensured, the main goal pursued from the practical point of view. Finally, we support the theoretical results with several numerical simulations.
Citation: Tomás Caraballo, Javier López-de-la-Cruz. Bounded random fluctuations on the input flow in chemostat models with wall growth and non-monotonic kinetics[J]. AIMS Mathematics, 2021, 6(4): 4025-4052. doi: 10.3934/math.2021239
This paper investigates a chemostat model with wall growth and Haldane consumption kinetics. In addition, bounded random fluctuations on the input flow, which are modeled by means of the well-known Ornstein-Uhlenbeck process, are considered to obtain a much more realistic model fitting in a better way the phenomena observed by practitioners in real devices. Once the existence and uniqueness of global positive solution has been proved, as well as the existence of deterministic absorbing and attracting sets, the random dynamics inside the attracting set is studied in detail to provide conditions under which persistence of species is ensured, the main goal pursued from the practical point of view. Finally, we support the theoretical results with several numerical simulations.

| [1] | J. Monod, La technique de culture continue: Théorie et applications, Annales de l'Institute Pasteur, 79 (1950), 390–410. |
| [2] | A. Novick, L. Szilard, Experiments with the chemostat on spontaneous mutations of bacteria, Proceedings of the National Academy of Sciences, 36 (1950), 708–719. |
| [3] | G. Stephanopoulos, R. Aris, A. Fredrickson, A stochastic analysis of the growth of competing microbial populations in a continuous biochemical reactor, Math. Biosci., 45 (1979), 99–135. |
| [4] | F. Stewart, B. Levin, The population biology of bacterial plasmids: A priori conditions for the existence of conjugationally transmitted factors, Genetics, 87 (1977), 209–228. |
| [5] | G. D'Ans, P. Kokotovic, D. Gottlieb, A nonlinear regulator problem for a model of biological waste treatment, IEEE T. Automat. Contr., 16 (1971), 341–347. |
| [6] | J. W. M. La Rivière, Microbial ecology of liquid waste treatment, In: Advances Microbial Ecology, Springer US, 1 (1977), 215–259. |
| [7] | R. Freter, Human Intestinal Microflora in Health and Disease, ch. Mechanisms that control the microflora in the large intestine, Academic Press, New York, 1983, 33–54. |
| [8] | R. Freter, An understanding of colonization of the large intestine requires mathematical analysis, Microecol. Therapy, 16 (1986), 147–155. |
| [9] | J. Barlow, F. de Noyelles, B. Peterson, J. Peterson, W. Schaffner, "Continuous flow nutrient bioassays with natural phytoplankton populations", G. Glass (Editor): Bioassay Techniques and Environmental Chemistry, John Wiley & Sons Ltd., 1973. |
| [10] | H. R. Bungay, M. L. Bungay, Microbial interactions in continuous culture, Advances Appl. Microbiol., 10 (1986), 269–290. |
| [11] |
I. F. Creed, D. M. McKnight, B. A. Pellerin, M. B. Green, B. A. Bergamaschi, G. R. Aiken et al., The river as a chemostat: Fresh perspectives on dissolved organic matter flowing down the river continuum, Canadian J. Fisheries Aquatic Sci., 72 (2015), 1272–1285. doi: 10.1139/cjfas-2014-0400

|
| [12] | A. Cunningham, R. M. Nisbet, Transients and oscillations in continuous cultures, Math. Microbiol., (1983), 77–103, 1983. |
| [13] | A. Fredrickson, G. Stephanopoulos, Microbial competition, Science, 213 (1981), 972–979. |
| [14] |
H. W. Jannasch, Steady state and the chemostat in ecology, Limnol. Oceanogr., 19 (1974), 716–720, 1974. doi: 10.4319/lo.1974.19.4.0716
|
| [15] |
J. Kalff, R. Knoechel, Phytoplankton and their dynamics in oligotrophic and eutrophic lakes, Annual Review Ecology Syst., 9 (1978), 475–495. doi: 10.1146/annurev.es.09.110178.002355
|
| [16] |
E. Rurangwa, M. C. J. Verdegem, Microorganisms in recirculating aquaculture systems and their management, Reviews Aquacult., 7 (2015), 117–130. doi: 10.1111/raq.12057
|
| [17] |
P. A. Taylor, J. L. Williams, Theoretical studies on the coexistence of competing species under continuous flow conditions, Cadandian J. Microbiol., 21 (1975), 90–98. doi: 10.1139/m75-013
|
| [18] |
H. Veldcamp, Ecological studies with the chemostat, Advances Microbial Ecol., 1 (1977), 59–95. doi: 10.1007/978-1-4615-8219-9_2
|
| [19] | P. Waltman, Competition Models in Population Biology. CBMS-NSF Regional Conference Series in Applied Mathematics, 1983, Society for Industrial and Applied Mathematics, Philadelphia. |
| [20] | P. Waltman, S. P. Hubbel, S. B. Hsu, Theoretical and experimental investigations of microbial competition in continuous culture, Modeling Differential Equations in Biol. (Conf., southern Illinois Univ. Carbonadle, III., 1978), 58 (1980), 107–152. |
| [21] | J. Harmand, C. Lobry, A. Rapaport, T. Sari, The Chemostat: Mathematical Theory of Micro-organisms Cultures. Wiley, Chemical Engineering Series, John Wiley & Sons, Inc., 2017. |
| [22] | V. Sree Hari Rao, P. Raja Sekhara Rao, Dynamic Models and Control of Biological Systems. Springer-Verlag, Heidelberg, 2009. |
| [23] | S. Pilyugin, P. Waltman, The simple chemostat with wall growth, Siam J. Appl. Math.-SIAMAM, 59 (1999), 09. |
| [24] | J. S. H. Haldane, Enzymes. Longmans Green and Co, London, 1930. |
| [25] |
J. Andrews, A mathematical model for the continuous culture of microorganisms utilizing inhibitory substrates, Biotechnol. Bioeng., 10 (1968), 707–723. doi: 10.1002/bit.260100602
|
| [26] | T. Caraballo, X. Han, Applied Nonautonomous and Random Dynamical Systems, Applied Dynamical Systems, Springer International Publishing, 2016. |
| [27] | T. Caraballo, X. Han, P. E. Kloeden, A. Rapaport, Continuous and Distributed Systems II, ch. Dynamics of Non autonomous Chemostat Models, Springer International Publishing, Cham, 103–120, 2015. |
| [28] | T. Caraballo, X. Han, P. E. Kloeden, Chemostats with time-dependent inputs and wall growth, Appl. Math. Inf. Sci., 9 (2015), 2283–2296. |
| [29] |
M. El Hajji, A. Rapaport, Practical coexistence of two species in the chemostat-a slow-fast characterization, Math. Biosci., 218 (2009), 33–39. doi: 10.1016/j.mbs.2008.12.003
|
| [30] | T. Caraballo, M. J. Garrido-Atienza, J. López-de-la-Cruz and A. Rapaport, Modeling and analysis of random and stochastic input flows in the chemostat model, Discrete Cont. Dyn-B, 24 (2018), 3591–3614. |
| [31] |
T. Caraballo, R. Colucci, J. López-de-la-Cruz and A. Rapaport, A way to model stochastic perturbations in population dynamics models with bounded realizations, Commun. Nonlinear Sci., 77 (2019), 239–257. doi: 10.1016/j.cnsns.2019.04.019
|
| [32] |
T. Caraballo, R. Colucci, J. López-de-la-Cruz, A. Rapaport, Study of the chemostat model with non-monotonic growth under random disturbances on the removal rate, Math. Biosci. Eng., 17 (2020), 7480–7501. doi: 10.3934/mbe.2020382
|
| [33] | T. Caraballo, J. López-de-la-Cruz, A. Rapaport, Modeling bounded random fluctuations in biological systems: Application to the chemostat model with two species, IFAC-PapersOnLine, 52 (2019), 187–192. |
| [34] |
J. López-de-la-Cruz, Random and stochastic disturbances on the input flow in chemostat models with wall growth, Stoch. Anal. Appl., 37 (2019), 668–698. doi: 10.1080/07362994.2019.1605911
|
| [35] | X. Li, X. Yang, T. Huang, Persistence of delayed cooperative models: Impulsive control method, Appl. Math. Comput., 342 (2019), 130–146. |
| [36] | X. Li, J. Shen, R. Rakkiyappan, Persistent impulsive effects on stability of functional differential equations with finite or infinite delay, Appl. Math. Comput., 329 (2018), 14–22. |
| [37] | H. L. Smith, P. Waltman, The theory of the chemostat: dynamics of microbial competition, Cambridge University Press, 1995. |
| [38] |
C. Xu, S. Yuan, An analogue of break-even concentration in a simple stochastic chemostat model, Appl. Math. Lett., 48 (2015), 62–68, 2015. doi: 10.1016/j.aml.2015.03.012
|
| [39] |
D. Zhao, S. Yuan, Critical result on the break-even concentration in a single-species stochastic chemostat model, J. Math. Anal. Appl., 434 (2016), 1336–1345. doi: 10.1016/j.jmaa.2015.09.070
|
| [40] |
T. Caraballo, P. E. Kloeden, B. Schmalfuss, Exponentially stable stationary solutions for stochastic evolution equations and their perturbation, Appl. Math. Optim., 50 (2004), 183–207. doi: 10.1007/s00245-004-0802-1
|
| [41] | L. Arnold, Random Dynamical Systems, Springer Berlin Heidelberg, 1998. |
| [42] | T. Caraballo, M. J. Garrido-Atienza, J. López-de-la-Cruz, Some Aspects Concerning the Dynamics of Stochastic Chemostats, Springer International Publishing, 69 (2016), 227–246. |
| [43] |
T. Caraballo, M. J. Garrido-Atienza, J. López-de-la-Cruz, Dynamics of some stochastic chemostat models with multiplicative noise, Commun. Pure Appl. Anal., 16 (2017), 1893–1914. doi: 10.3934/cpaa.2017092
|
| [44] | G. Bastin, D. Dochain, On-line estimation and adaptive control of bioreactors, Elsevier, 1990. |
| [45] |
A. Rapaport, J. Harmand, Robust regulation of a class of partially observed nonlinear continuous bioreactors, J. Process Contr., 12 (2002), 291–302. doi: 10.1016/S0959-1524(01)00029-4
|
| [46] |
B. Satishkumar, M. Chidambaram, Control of unstable bioreactor using fuzzy tuned PI controller, Bioprocess Eng., 20 (1999), 127. doi: 10.1007/s004490050570
|
| [47] |
A. Schaum, J. Alvarez, T. Lopez-Arenas, Saturated PI control of continuous bioreactors with haldane kinetics, Chemical Eng. Sci., 68 (2012), 520–529. doi: 10.1016/j.ces.2011.10.006
|
| [48] | A. Rapaport, I. Haidar, J. Harmand, Global dynamics of the buffered chemostat for a general class of response functions, J. Math. Biol., 71 (2014), 69–98. |
| [49] |
A. Rapaport, J. Harmand, Biological control of the chemostat with nonmonotonic response and different removal rates, Math. Biosci. Eng., 5 (2008), 539–547. doi: 10.3934/mbe.2008.5.539
|
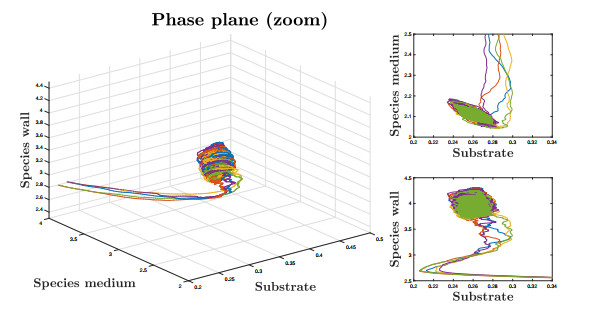
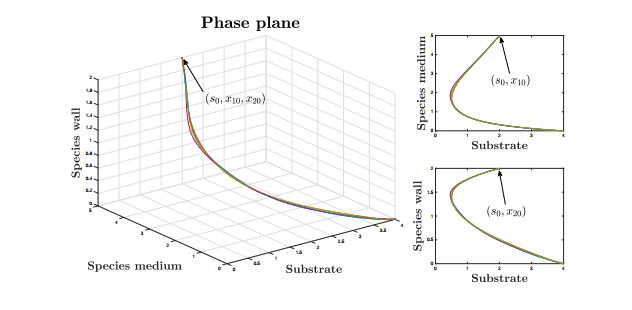
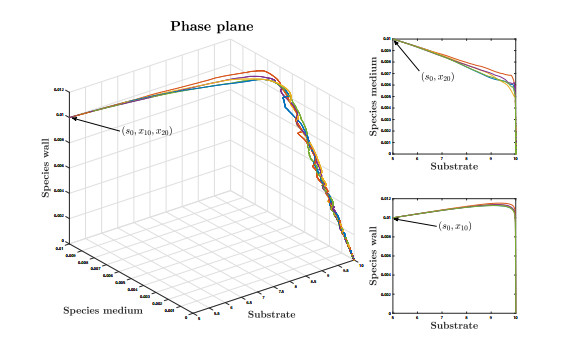
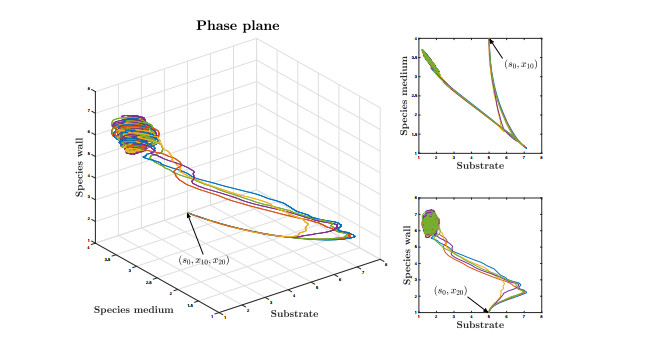
Figures(9)

Tomás Caraballo, Javier López-de-la-Cruz. Bounded random fluctuations on the input flow in chemostat models with wall growth and non-monotonic kinetics[J]. AIMS Mathematics, 2021, 6(4): 4025-4052. doi: 10.3934/math.2021239







 DownLoad:
DownLoad: