Citation: Silvia Colodrero. Conjugated polymers as functional hole selective layers in efficient metal halide perovskite solar cells[J]. AIMS Materials Science, 2017, 4(4): 956-969. doi: 10.3934/matersci.2017.4.956

| [1] |
Sum TC, Mathews N (2014) Advancements in perovskite solar cells: photophysics behind the photovoltaics. Energ Environ Sci 7: 2518–2534. doi: 10.1039/C4EE00673A

|
| [2] |
Stoumpos CC, Kanatzidis MG (2015) The renaissance of halide perovskites and their evolution as emerging semiconductors. Accounts Chem Res 48: 2791–2802. doi: 10.1021/acs.accounts.5b00229
|
| [3] |
Chen Q, De Marco N, Yang Y, et al. (2015) Under the spotlight: The organic-inorganic hybrid halide perovskite for optoelectronic applications. Nano Today 10: 355–396. doi: 10.1016/j.nantod.2015.04.009
|
| [4] |
Zhao Y, Zhu K (2016) Organic-inorganic hybrid lead halide perovskites for optoelectronic and electronic applications. Chem Soc Rev 45: 655–689. doi: 10.1039/C4CS00458B
|
| [5] |
Kojima A, Teshima K, Shirai Y, et al. (2009) Organometal halide perovskites as visible-light sensitizers for photovoltaic cells. J Am Chem Soc 131: 6050–6051. doi: 10.1021/ja809598r
|
| [6] |
Kim HS, Lee CR, Im JH, et al. (2012) Lead iodide perovskite sensitized all-solid-state submicron thin film mesoscopic solar cell with efficiency exceeding 9%. Sci Rep 2: 591. doi: 10.1038/srep00591
|
| [7] |
Lee MM, Teuscher J, Miyasaka T, et al. (2012) Efficient hybrid solar cells based on meso-superstructured organometal halide perovskites. Science 338: 643–647. doi: 10.1126/science.1228604
|
| [8] |
Saliba M, Matsui T, Domanski K, et al. (2016) Incorporation of rubidium cations into perovskite solar cells improves photovoltaic performance. Science 354: 206–209. doi: 10.1126/science.aah5557
|
| [9] | Green MA, Emery K, Hishikawa Y, et al. (2014) Solar cell efficiency tables (Version 45). Prog Photovolt Res Appl 23: 1–9. |
| [10] |
Burschka J, Pellet N, Moon SJ, et al. (2013) Sequential deposition as a route to high-performance perovskite-sensitized solar cells. Nature 499: 316–319. doi: 10.1038/nature12340
|
| [11] |
Eperon GE, Burlakov VM, Docampo P, et al. (2014) Morphological control for high performance, solution-processed planar heterojunction perovskite solar cells. Adv Funct Mater 24: 151–157. doi: 10.1002/adfm.201302090
|
| [12] |
Liang PW, Liao CY, Chueh CC, et al. (2014) Additive enhanced crystallization of solution-processed perovskite for highly efficient planar-heterojunction solar cells. Adv Mater 26: 3748–3754. doi: 10.1002/adma.201400231
|
| [13] |
Xie FX, Zhang D, Su H, et al. (2015) Vacuum-assisted thermal annealing of CH3NH3PbI3 for highly stable and efficient perovskite solar cells. ACS Nano 9: 639–646. doi: 10.1021/nn505978r
|
| [14] |
Zuo L, Gu Z, Ye T, et al. (2015) Enhanced photovoltaic performance of CH3NH3PbI3 perovskite solar cells through interfacial engineering using self-assembling monolayer. J Am Chem Soc 137: 2674–2679. doi: 10.1021/ja512518r
|
| [15] |
Seo J, Noh JH, Seok Sl (2016) Rational strategies for efficient perovskite solar cells. Accounts Chem Res 49: 562–572. doi: 10.1021/acs.accounts.5b00444
|
| [16] |
Eperon GE, Stranks SD, Menelaou C, et al. (2014) Formamidinium lead trihalide: a broadly tunable perovskite for efficient planar heterojunction solar cells. Energ Environ Sci 7: 982–988. doi: 10.1039/c3ee43822h
|
| [17] |
Xiao Z, Dong Q, Bi C, et al. (2014) Solvent annealing of perovskite-induced crystal growth for photovoltaic-device efficiency enhancement. Adv Mater 26: 6503–6509. doi: 10.1002/adma.201401685
|
| [18] |
Long M, Zhang T, Xu W, et al. (2017) Large-grain formamidinium PbI3−xBrx for high-performance perovskite solar cells via intermediate halide exchange. Adv Energy Mater 7: 1601882. doi: 10.1002/aenm.201601882
|
| [19] |
Long M, Zhang T, Zhu H, et al. (2017) Textured CH3NH3PbI3 thin film with enhanced stability for high performance perovskite solar cells. Nano Energy 33: 485–496. doi: 10.1016/j.nanoen.2017.02.002
|
| [20] |
Williams ST, Rajagopal A, Chueh CC, et al. (2016) Current challenges and prospective research for upscaling hybrid perovskite photovoltaics. J Phys Chem Lett 7: 811–819. doi: 10.1021/acs.jpclett.5b02651
|
| [21] |
Stranks SD, Eperon GE, Grancini G, et al. (2013) Electron-hole diffusion lengths exceeding 1 micrometer in an organometal trihalide perovskite absorber. Science 342: 341–344. doi: 10.1126/science.1243982
|
| [22] |
Xing G, Mathews N, Sun S, et al. (2013) Long-range balanced electron- and hole-transport lengths in organic-inorganic CH3NH3PbI3. Science 342: 344–347. doi: 10.1126/science.1243167
|
| [23] |
Shi D, Adinolfi V, Comin R, et al. (2015) Low trap-state density and long carrier diffusion in organolead trihalide perovskite single crystals. Science 347: 519–522. doi: 10.1126/science.aaa2725
|
| [24] |
De Quilettes DW, Vorpahl SM, Stranks SD, et al. (2015) Impact of microstructure on local carrier lifetime in perovskite solar cells. Science 348: 683–686. doi: 10.1126/science.aaa5333
|
| [25] |
Yang WS, Noh JH, Jeon NJ, et al. (2015) High-performance photovoltaic perovskite layers fabricated through intramolecular exchange. Science 348: 1234–1237. doi: 10.1126/science.aaa9272
|
| [26] |
Jeon NJ, Noh JH, Yang WS, et al. (2015) Compositional engineering of perovskite materials for high-performance solar cells. Nature 517: 476–480. doi: 10.1038/nature14133
|
| [27] |
Ke W, Fang G, Liu Q, et al. (2015) Low-temperature solution-processed tin oxide as an alternative electron transporting layer for efficient perovskite solar cells. J Am Chem Soc 137: 6730–6733. doi: 10.1021/jacs.5b01994
|
| [28] |
Anaraki EH, Kermanpur A, Steier L, et al. (2016) Highly efficient and stable planar perovskite solar cells by solution-processed tin oxide. Energ Environ Sci 9: 3128–3134. doi: 10.1039/C6EE02390H
|
| [29] |
Xu X, Zhang H, Shi J, et al. (2015) Highly efficient planar perovskite solar cells with a TiO2/ZnO electron transport bilayer. J Mater Chem A 3: 19288–19293. doi: 10.1039/C5TA04239A
|
| [30] |
Okamoto Y, Suzuki Y (2016) Mesoporous BaTiO3/TiO2 double layer for electron transport in perovskite solar cells. J Phys Chem C 120: 13995–14000. doi: 10.1021/acs.jpcc.6b04642
|
| [31] |
Nguyen WH, Bailie CD, Unger EL, et al. (2014) Enhancing the hole-conductivity of spiro-OMeTAD without oxygen or lithium salts by using spiro(TFSI)2 in perovskite and dye-sensitized solar cells. J Am Chem Soc 136: 10996–11001. doi: 10.1021/ja504539w
|
| [32] |
Kim HS, Seo JY, Park NG (2016) Impact of selective contacts on long-term stability of CH3NH3PbI3 perovskite solar cells. J Phys Chem C 120: 27840–27848. doi: 10.1021/acs.jpcc.6b09412
|
| [33] |
Liu Y, Chen Q, Duan HS, et al. (2015) A dopant-free organic hole transport material for efficient planar heterojunction perovskite solar cells. J Mater Chem A 3: 11940–11947. doi: 10.1039/C5TA02502H
|
| [34] |
Xu J, Voznyy O, Comin R, et al. (2016) Crosslinked remote-doped hole-extracting contacts enhance stability under accelerated lifetime testing in perovskite solar cells. Adv Mater 28: 2807–2815. doi: 10.1002/adma.201505630
|
| [35] |
Kim GW, Kang G, Kim J, et al. (2016) Dopant-free polymeric hole transport materials for highly efficient and stable perovskite solar cells. Energ Environ Sci 9: 2326–2333. doi: 10.1039/C6EE00709K
|
| [36] |
Habisreutinger SN, McMeekin DP, Snaith HJ, et al. (2016) Research Update: Strategies for improving the stability of perovskite solar cells. APL Mater 4: 091503. doi: 10.1063/1.4961210
|
| [37] |
Yang J, Siempelkamp BD, Liu D, et al. (2015) Investigation of CH3NH3PbI3 degradation rates and mechanisms in controlled humidity environments using in situ techniques. ACS Nano 9: 1955–1963. doi: 10.1021/nn506864k
|
| [38] |
Christians JA, Miranda Herrera PA, Kamat PV (2015) Transformation of the excited state and photovoltaic efficiency of CH3NH3PbI3 perovskite upon controlled exposure to humidified air. J Am Chem Soc 137: 1530–1538. doi: 10.1021/ja511132a
|
| [39] |
Leguy AMA, Hu Y, Campoy-Quiles M, et al. (2015) Reversible Hydration of CH3NH3PbI3 in Films, Single Crystals, and Solar Cells. Chem Mater 27: 3397–3407. doi: 10.1021/acs.chemmater.5b00660
|
| [40] |
Habisreutinger SN, Leijtens T, Eperon GE, et al. (2014) Carbon nanotube/polymer composites as a highly stable hole collection layer in perovskite solar cells. Nano Lett 14: 5561–5568. doi: 10.1021/nl501982b
|
| [41] |
Smith IC, Hoke ET, Solis-Ibarra D, et al. (2014) A layered hybrid perovskite solar-cell absorber with enhanced moisture stability. Angew Chem 126: 11414–11417. doi: 10.1002/ange.201406466
|
| [42] |
Leijtens T, Giovenzana T, Habisreutinger SN, et al. (2016) Hydrophobic organic hole transporters for improved moisture resistance in metal halide perovskite solar cells. ACS Appl Mater Interfaces 8: 5981–5989. doi: 10.1021/acsami.5b10093
|
| [43] |
Hwang I, Jeong I, Lee J, et al. (2015) Enhancing stability of perovskite solar cells to moisture by the facile hydrophobic passivation. ACS Appl Mater Interfaces 7: 17330–17336. doi: 10.1021/acsami.5b04490
|
| [44] |
Sirringhaus H, Tessler N, Friend RH (1998) Integrated optoelectronic devices based on conjugated polymers. Science 280: 1741–1744. doi: 10.1126/science.280.5370.1741
|
| [45] |
Pei Q, Yu G, Zhang C, et al. (1995) Polymer light-emitting electrochemical cells. Science 269: 1086–1088. doi: 10.1126/science.269.5227.1086
|
| [46] |
Yu G, Gao J, Hummelen JC, et al. (1995) Polymer photovoltaic cells: enhanced efficiencies via a network of internal donor–acceptor heterojunctions. Science 270: 1789–1791. doi: 10.1126/science.270.5243.1789
|
| [47] |
Forrest SR (2004) The path to ubiquitous and low-cost organic electronic appliances on plastic. Nature 428: 911–918. doi: 10.1038/nature02498
|
| [48] |
Bi D, Yang L, Boschloo G, et al. (2013) Effect of different hole transport materials on recombination in CH3NH3PbI3 perovskite-sensitized mesoscopic solar cells. J Phys Chem Lett 4: 1532–1536. doi: 10.1021/jz400638x
|
| [49] |
Heo JH, Im SH, Noh JH, et al. (2013) Efficient inorganic-organic hybrid heterojunction solar cells containing perovskite compound and polymeric hole conductors. Nat Photonics 7: 486–491. doi: 10.1038/nphoton.2013.80
|
| [50] |
Kwon YS, Lim J, Yun HJ, et al. (2014) A diketopyrrolopyrrole-containing hole transporting conjugated polymer for use in efficient stable organic-inorganic hybrid solar cells based on a perovskite. Energ Environ Sci 7: 1454–1460. doi: 10.1039/c3ee44174a
|
| [51] |
Zhu Z, Bai Y, Lee HK, et al. (2014) Polyfluorene derivatives are high-performance organic hole-transporting materials for inorganic-organic hybrid perovskite solar cells. Adv Funct Mater 24: 7357–7365. doi: 10.1002/adfm.201401557
|
| [52] |
Kranthiraja K, Gunasekar K, Kim H, et al. (2017) High-performance long-term-stable dopant-free perovskite solar cells and additive-free organic solar cells by employing newly designed multirole π-conjugated polymers. Adv Mater 29: 1700183. doi: 10.1002/adma.201700183
|
| [53] |
Long M, Chen Z, Zhang T, et al. (2016) Ultrathin efficient perovskite solar cells employing a periodic structure of a composite hole conductor for elevated plasmonic light harvesting and hole collection. Nanoscale 8: 6290–6299. doi: 10.1039/C5NR05042A
|
| [54] |
He Z, Xiao B, Liu F, et al. (2015) Single-junction polymer solar cells with high efficiency and photovoltage. Nat Photonics 9: 174–179. doi: 10.1038/nphoton.2015.6
|
| [55] |
Chen D, Nakahara A, Wei D, et al. (2011) P3HT/PCBM bulk heterojunction organic photovoltaics: correlating efficiency and morphology. Nano Lett 11: 561–567. doi: 10.1021/nl103482n
|
| [56] |
Di Giacomo F, Razza S, Matteocci F, et al. (2014) High efficiency CH3NH3PbI(3−x)Clx, perovskite solar cells with poly(3-hexylthiophene) hole transport layer. J Power Sources 251: 152–156. doi: 10.1016/j.jpowsour.2013.11.053
|
| [57] |
Xiao JY, Shi JJ, Liu HB, et al. (2015) Efficient CH3NH3PbI3 perovskite solar cells based on graphdiyne (GD)-modified P3HT hole-transporting material. Adv Energy Mater 5: 1401943. doi: 10.1002/aenm.201401943
|
| [58] |
Guo YL, Liu C, Inoue K, et al. (2014) Enhancement in the efficiency of an organic-inorganic hybrid solar cell with a doped P3HT hole-transporting layer on a void-free perovskite active layer. J Mater Chem A 2: 13827–13830. doi: 10.1039/C4TA02976C
|
| [59] |
Zhao N, Osedach TP, Chang LY, et al. (2010) Colloidal PbS quantum dot solar cells with high fill factor. ACS Nano 4: 3743–3752. doi: 10.1021/nn100129j
|
| [60] |
Koster LJA, Mihailetchi VD, Ramaker R, et al. (2005) Light intensity dependence of open-circuit voltage of polymer: fullerene solar cells. Appl Phys Lett 86: 123509–123509. doi: 10.1063/1.1889240
|
| [61] |
Mandoc M, Kooistra F, Hummelen J, et al. (2007) Effect of traps on the performance of bulk heterojunction organic solar cells. Appl Phys Lett 91: 263505. doi: 10.1063/1.2821368
|
| [62] |
Wetzelaer GAH, Scheepers M, Sempere AM, et al. (2015) Trap-assisted non-radiative recombination in organic-inorganic perovskite solar cells. Adv Mater 27: 1837–1841. doi: 10.1002/adma.201405372
|
| [63] |
Sherkar TS, Momblona C, Gil-Escrig L, et al. (2017) Improving the performance of perovskite solar cells: insights from a validated device model. Adv Energy Mater 7: 1602432. doi: 10.1002/aenm.201602432
|
| [64] |
Sherkar TS, Momblona, C, Gil-Escrig L, et al. (2017) Recombination in perovskite solar cells: significance of grain boundaries, interface traps, and defect ions. ACS Energy Lett 2: 1214–1222. doi: 10.1021/acsenergylett.7b00236
|
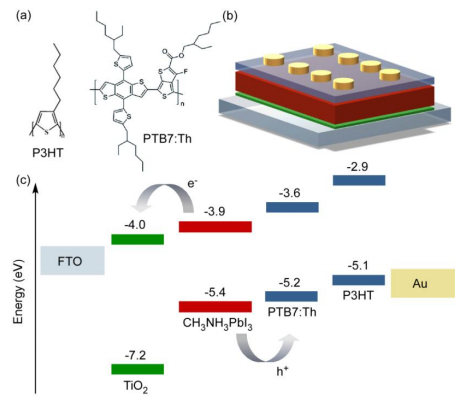
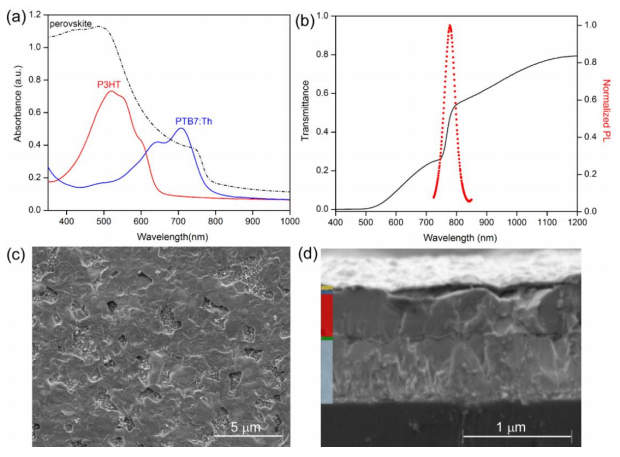
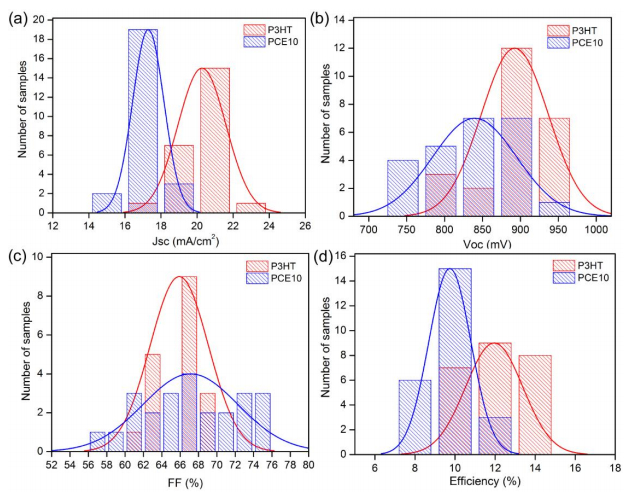
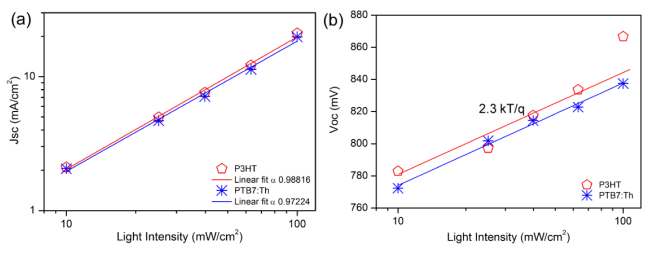
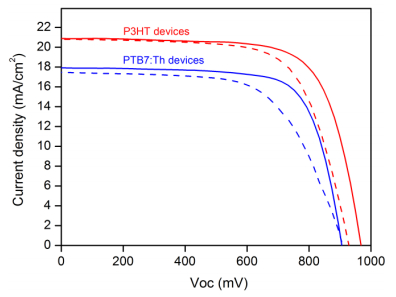
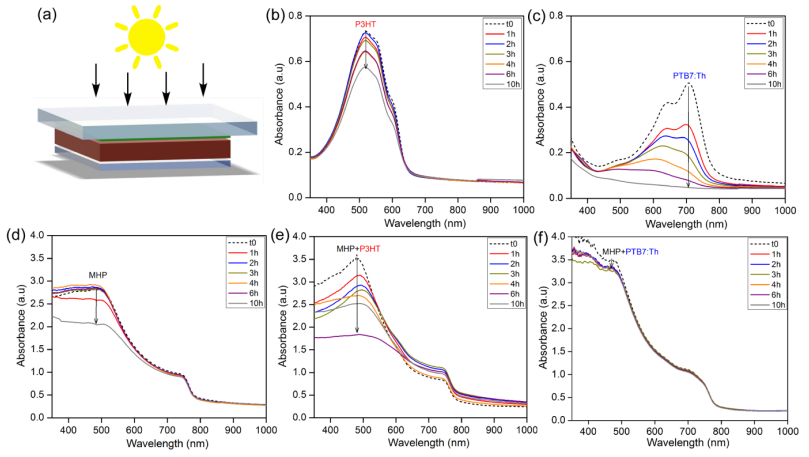
Figures(6) / Tables(1)

Silvia Colodrero. Conjugated polymers as functional hole selective layers in efficient metal halide perovskite solar cells[J]. AIMS Materials Science, 2017, 4(4): 956-969. doi: 10.3934/matersci.2017.4.956







 DownLoad:
DownLoad: