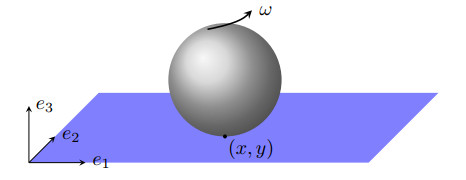
We propose a numerical scheme for the time-integration of nonholonomic mechanical systems, both conservative and nonconservative. The scheme is obtained by simultaneously discretizing the constraint equations and the Herglotz variational principle. We validate the method using numerical simulations and contrast them against the results of standard methods from the literature.
Citation: Elias Maciel, Inocencio Ortiz, Christian E. Schaerer. A Herglotz-based integrator for nonholonomic mechanical systems[J]. Journal of Geometric Mechanics, 2023, 15(1): 287-318. doi: 10.3934/jgm.2023012
We propose a numerical scheme for the time-integration of nonholonomic mechanical systems, both conservative and nonconservative. The scheme is obtained by simultaneously discretizing the constraint equations and the Herglotz variational principle. We validate the method using numerical simulations and contrast them against the results of standard methods from the literature.

| [1] | E. Hairer, C. Lubich, G. Wanner, Solving Geometric Numerical Integration: Structure-Preserving Algorithms, 2006. |
| [2] |
J. M. Wendlandt, J. E Marsden, Mechanical integrators derived from a discrete variational principle, Physica. D., 106 (1997), 223–246. https://doi.org/10.1016/S0167-2789(97)00051-1 doi: 10.1016/S0167-2789(97)00051-1

|
| [3] |
J. E. Marsden, M. West, Discrete mechanics and variational integrators, Acta. Numer., 10 (2001), 357–514. https://doi.org/10.1017/S096249290100006X doi: 10.1017/S096249290100006X
|
| [4] |
J. Y. Ding, Z. K. Pan, Higher order variational integrators for multibody system dynamics with constraints, Adv. Mech. Eng., 6 (2014), 383680. https://doi.org/10.1155/2014/383680 doi: 10.1155/2014/383680
|
| [5] |
S. Leyendecker, J. E. Marsden, M. Ortiz, Variational integrators for constrained dynamical systems, ZAMM, 88 (2008), 677–708. https://doi.org/10.1002/zamm.200700173 doi: 10.1002/zamm.200700173
|
| [6] | A. Stern, Y. Y. Tong, M. Desbrun, J. E. Marsden, Geometric computational electrodynamics with variational integrators and discrete differential forms, In Geometry, mechanics, and dynamics, Springer, (2015), 437–475. |
| [7] |
S. Ober-Blöbaum, M. Tao, M. L. Cheng, H. Owhadi, J. E. Marsden, Variational integrators for electric circuits, J. Comput. Phys., ZAMM, 242 (2013), 498–530. https://doi.org/10.1002/zamm.200700173 doi: 10.1002/zamm.200700173
|
| [8] |
S. Ober-Blöbaum, Galerkin variational integrators and modified symplectic Runge-Kutta methods, Ima. J. Numer. Anal., 37 (2016), 375–406. https://doi.org/10.1093/imanum/drv062 doi: 10.1093/imanum/drv062
|
| [9] |
J. E. Marsden, S. Pekarsky, S. Shkoller, M. West, Variational methods, multisymplectic geometry and continuum mechanics, J. Geom. Phys., 38 (2001), 253–284. https://doi.org/10.1016/S0393-0440(00)00066-8 doi: 10.1016/S0393-0440(00)00066-8
|
| [10] |
T. Y. Lee, M. Leok, N. H. McClamroch, Lie group variational integrators for the full body problem in orbital mechanics, Celest. Mech. Dyn. Astr., 98 (2007), 121–144. https://doi.org/10.1007/s10569-007-9073-x doi: 10.1007/s10569-007-9073-x
|
| [11] | X. F. Shen, M. Leok, Lie group variational integrators for rigid body problems using quaternions, arXiv preprint arXiv: 1705.04404, 2017. |
| [12] |
F. Demoures, F. Gay-Balmaz, S. Leyendecker, S. Ober-Blöbaum, T. S. Ratiu, Y. Weinand, Discrete variational lie group formulation of geometrically exact beam dynamics, Numer. Math., 130 (2015), 73–123. https://doi.org/10.1007/s00211-014-0659-4 doi: 10.1007/s00211-014-0659-4
|
| [13] | J. E. Marsden, J. M, Wendlandt, Mechanical systems with symmetry, variational principles, and integration algorithms, In Current and future directions in applied mathematics, Springer, (1997), 219–261. https://doi.org/10.1007/978-1-4612-2012-1-18 |
| [14] |
T. Lee, M. Leok, N. H. McClamroch, Lie group variaional integrator for the full body problem in orbital mechanics, Celest. Mech. Dyn. Astr., 98 (2007), 121–144. https://doi.org/10.1007/s10569-007-9073-x doi: 10.1007/s10569-007-9073-x
|
| [15] | L. Colombo, F. Jiménez, D. M. de Diego. Variational integrators for underactuated mechanical control systems with symmetries, arXiv preprint arXiv: 1209.6315, 2012. |
| [16] | M. Leok, An overview of lie group variational integrators and their applications to optimal control, In International conference on scientific computation and differential equations, The French National Institute for Research in Computer Science and Control, 2007. |
| [17] | C. M. Campos, S. Ober-Blöbaum, E. Trélat, High order variational integrators in the optimal control of mechanical systems, arXiv preprint arXiv: 1502.00325, 2015. |
| [18] | I. I. Hussein, M. Leok, A. K. Sanyal, A. M. Bloch, A discrete variational integrator for optimal control problems on so (3), In Proceedings of the 45th IEEE Conference on Decision and Control, IEEE, (2006), 6636–6641. |
| [19] |
M. de León, D. M. de Diego, A. Santamaría-Merino, Discrete variational integrators and optimal control theory, Adv. Comput. Math., 26 (2007), 251–268. https://doi.org/10.1007/s10444-004-4093-5 doi: 10.1007/s10444-004-4093-5
|
| [20] |
A. Lew, J. E. Marsden, M. Ortiz, M. West, Asynchronous variational integrators, Arch. Ration. Mech. An., 167 (2003), 85–146. https://doi.org/10.1007/s00205-002-0212-y doi: 10.1007/s00205-002-0212-y
|
| [21] |
J. Hall, M. Leok, Spectral variational integrators, Numer. Math., 130 (2015), 681–740. https://doi.org/10.1007/s00211-014-0679-0 doi: 10.1007/s00211-014-0679-0
|
| [22] |
M. Leok, T. Shingel, General techniques for constructing variational integrators, Front. Math. China., 7 (2012), 273–303. https://doi.org/10.1007/s11464-012-0190-9 doi: 10.1007/s11464-012-0190-9
|
| [23] |
D. JN. Limebeer, S. Ober-Blöbaum, F. Haddad. Farshi, Variational integrators for dissipative systems, Ieee. T. Automat. Contr., 65 (2020), 1381–1396. https://doi.org/10.1109/TAC.2020.2965059 doi: 10.1109/TAC.2020.2965059
|
| [24] |
Z. Manchester, N. Doshi, R. J. Wood, S. Kuindersma, Contact-implicit trajectory optimization using variational integrators, Ind. Robot., 38 (2019), 1463–1476. https://doi.org/10.1177/0278364919849235 doi: 10.1177/0278364919849235
|
| [25] |
V. Duruisseaux, J. Schmitt, M. Leok, Adaptive hamiltonian variational integrators and applications to symplectic accelerated optimization, Siam. J. Sci. Comput., 43 (2021), A2949–A2980. https://doi.org/10.1137/20M1383835 doi: 10.1137/20M1383835
|
| [26] | S. M. Man, Q. Gao, W. X. Zhong, Variational integrators in holonomic mechanics, Mathematics, 8 (2020). ttps: //doi.org/10.3390/math8081358 |
| [27] |
B. Tran, M. Leok, Multisymplectic hamiltonian variational integrators, Int. J. Comput. Math., 99 (2022), 113–157. https://doi.org/10.1080/00207160.2021.1999427 doi: 10.1080/00207160.2021.1999427
|
| [28] |
M. Leok, Variational discretizations of gauge field theories using group-equivariant interpolation, Found. Comput. Math., 19 (2019), 965–989. https://doi.org/10.1007/s10208-019-09420-4 doi: 10.1007/s10208-019-09420-4
|
| [29] | L. Colombo, M. G. Fernández, D. M. de Diego, Variational integrators for non-autonomous systems with applications to stabilization of multi-agent formations, arXiv preprint arXiv: 2202.01471, 2022. |
| [30] |
L. J. Colombo, H. G. de Marina, A variational integrator for the distance-based formation control of multi-agent systems, IFAC-PapersOnLine, 51 (2018), 76–81. https://doi.org/10.1016/j.ifacol.2018.12.014 doi: 10.1016/j.ifacol.2018.12.014
|
| [31] | A. Lew, J. E. Marsden, M. Ortiz, M. West, An overview of variational integrators, International Center for Numerical Methods in Engineering (CIMNE), 2004. https://doi.org/10.1002/nme.958 |
| [32] | A. Lew, J. E. Marsden, M. Ortiz, Variational time integrators, Int. J. Numer. Meth. Eng., 60 (2004), 153–212. |
| [33] | A. J. Lew, P. Mata A, A Brief Introduction to Variational Integrators, Springer International Publishing, Cham, 2016. |
| [34] | E. Maciel, I. Ortiz, C. E. Schaerer, A comparison of geometric integrators for nonconservative and nonholonomic systems, In Proceedings of the Ibero-Latin-American Congress on Computational Methods in Engineering. Universidade Federal da Integração Latino-Americana, 2020. |
| [35] | J. Cortés, S. Martínez, Non-holonomic integrators, Nonlinearity, 14 (2001), 1365–1392. https://doi.org/10.1088/0951-7715/14/5/322 |
| [36] |
M. Vermeeren, A. Bravetti, M. Seri, Contact variational integrators, J Phys. A-Math. Theor., 44 (2019), 445206. https://doi.org/10.1088/1751-8121/ab4767 doi: 10.1088/1751-8121/ab4767
|
| [37] | D. Holm, Geometric Mechanics - Part I: Dynamics and Symmetry, World Scientific Publishing Company, 2011. https://doi.org/10.1142/p801 |
| [38] | D. Holm, Geometric Mechanics - Part II: Rotating, Translating and Rolling, World Scientific Publishing Company, 2011. https://doi.org/10.1142/p802 |
| [39] | D. Holm, Geometric Mechanics and Symmetry: from finite o infinite dimensions, Oxford University Press, 2009. |
| [40] | R. Abraham, J. Marsden, Foundations of Mechanics. American Mathematical Society, 2008. https://doi.org/10.1090/chel/364 |
| [41] | J. E. Marsden, T. S. Ratiu, Introduction to Mechanics and Symmetry: a basic exposition of classical mechanical systems, Springer-Verlag New York, 2 edition, 1999. https://doi.org/10.1007/978-0-387-21792-5 |
| [42] | A. M. Bloch, J. E. Marsden, P. E. Crouch, J. Baillieul. Nonholonomic Mechanics and Control. Springer, 2015. https://doi.org/10.1007/978-1-4939-3017-3-54 |
| [43] | J. Cortés. Geometric, Control and Numerical Aspects of Nonholonomic Systems, Springer, 2002. |
| [44] | A. D. Lewis, Lagrangian mechanics, dynamics, and control, Preprint available online at http://penelope.mast.queensu.ca/~andrew, 237, 2003. |
| [45] |
M. de León, D. M. de Diego. On the geometry of non-holonomic lagrangian systems, J. Math. Phys., 37 (1996), 3389–3414. https://doi.org/10.1063/1.531571 doi: 10.1063/1.531571
|
| [46] |
J. F. Carinena, M. F. Ranada, Lagrangian systems with constraints: a geometric approach to the method of lagrange multipliers, J. Phys. A-math. Gen., 26 (1993), 1335. https://doi.org/10.1088/0305-4470/26/6/016 doi: 10.1088/0305-4470/26/6/016
|
| [47] |
O. Krupková, Mechanical systems with nonholonomic constraints, J. Math. Phys., 38 (1997), 5098–5126. https://doi.org/10.1063/1.532196 doi: 10.1063/1.532196
|
| [48] |
O. Krupková, The nonholonomic variational principle, J. Phys. A-math. Gen., 42 (2009), 185201. https://doi.org/10.1088/1751-8113/42/18/185201 doi: 10.1088/1751-8113/42/18/185201
|
| [49] | O. Krupková, Geometric mechanics on nonholonomic submanifolds, Commun. Math., 18 (2010), 51–77. |
| [50] |
M. de León, J. C. Marrero, D. M. de Diego, Mechanical systems with nonlinear constraints, Int. J. Theor. Phys., 36 (1997), 979–995. https://doi.org/10.1007/BF02435796 doi: 10.1007/BF02435796
|
| [51] | M. F. Rañada, Time-dependent lagrangian systems: A geometric approach to the theory of systems with constraints, J. Math. Phys., 35 (1994). https://doi.org/10.1063/1.530664 |
| [52] | M. de León, J. C. Marrero, D. M. De Diego. New Developments in Differential Geometry, Budapest 1996, chapter Time-Dependent Mechanical Systems With Non-Linear Constraints, Springer, Dordrecht, 1999. |
| [53] | B. Georgieva, The Variational Principle of Herglotz and Reletad Results, Geom. Integr. Quantiza., 12 (2011), 214–225. |
| [54] |
Y. Zhang, X. Tian, Conservation laws of nonconservative nonholonomic system based on Herglotz variational problem, Phys. Lett. A., 383 (2019), 691–696. https://doi.org/10.1016/j.physleta.2018.11.034 doi: 10.1016/j.physleta.2018.11.034
|
| [55] | M. de León, V. M. Jiménez, M. L. Valcázar, Contact hamiltonian systems with nonholonomic constraints, arXiv preprint arXiv: 1911.05409, 2019. |
| [56] |
A, Bravetti, Contact hamiltonian dynamics: the concept and its use, Entropy, 19 (2017), 535. https://doi.org/10.3390/e19100535 doi: 10.3390/e19100535
|
| [57] | A. Bravetti, Contact geometry and thermodynamics, Int J. Geom. Methods. M., 16 (2019). https://doi.org/10.1142/S0219887819400036 |
| [58] |
A. Bravetti, C.S. Lopez-Monsalvo, F. Nettel, Contact symmetries and hamiltonian thermodynamics, Ann. Phys-New. York., 361 (2015), 377–400. https://doi.org/10.1016/j.aop.2015.07.010 doi: 10.1016/j.aop.2015.07.010
|
| [59] |
A. Bravetti, C. Hans, D. Tapias, Contact hamiltonian mechanics, Ann. Phys-New. York., 376 (2017), 17–89. https://doi.org/10.1016/j.aop.2016.11.003 doi: 10.1016/j.aop.2016.11.003
|
| [60] |
M. J. Lazo, J. Paiva, J. TS. Amaral, G. SF. Frederico, An action principle for action-dependent lagrangians: Toward an action principle to non-conservative systems, J. Math. Phys., 59 (2018), 032902. https://doi.org/10.1063/1.50199364 doi: 10.1063/1.50199364
|
| [61] | M. de León, M. Lainz, A review on contact hamiltonian and lagrangian systems, arXiv preprint arXiv: 2011.05579, 2020. |
| [62] | A. López-Gordón, L. Colombo, M. de León, Nonsmooth herglotz variational principle, arXiv preprint arXiv: 2208.02033, 2022. |
| [63] |
A. Anahory Simoes, D. M. de Diego, M. L. Valcázar, M. de. León, On the geometry of discrete contact mechanics, J. Nonlinear. Sci., 31 (2021), 1–30. https://doi.org/10.1007/s00332-021-09708-2 doi: 10.1007/s00332-021-09708-2
|
| [64] | K. Modin, O. Verdier, What makes nonholonomic integrators work? Numer. Math., 145 (2020), 405–435. https://doi.org/10.1007/s00211-020-01126-y |
| [65] |
P. C. Paris, Li Zhang, A disk rolling on a horizontal surface without slip, Math. Comput. Model., 36 (2002), 855–860. https://doi.org/10.1016/S0895-7177(02)00232-7 doi: 10.1016/S0895-7177(02)00232-7
|
| [66] | E. Maciel, Propuesta de integrador geométrico para sistemas mecánicos no conservativos y no holonómicos, Master's thesis, Polytechnic School - National University of Asuncion, May 2021. |
 jgm-15-01-012-s001.pdf jgm-15-01-012-s001.pdf |
 |



















Figures(20) / Tables(1)
Elias Maciel, Inocencio Ortiz, Christian E. Schaerer. A Herglotz-based integrator for nonholonomic mechanical systems[J]. Journal of Geometric Mechanics, 2023, 15(1): 287-318. doi: 10.3934/jgm.2023012







 DownLoad:
DownLoad: