Citation: Yekaterina Zvorykina, Anna Tvorogova, Aleena Gladkikh, Ivan Vorobjev. Non-centrosomal MTs play a crucial role in organization of MT array in interphase fibroblasts[J]. AIMS Genetics, 2018, 5(2): 141-160. doi: 10.3934/genet.2018.2.141

| [1] |
Rodionov V, Nadezhdina E, Borisy G (1999) Centrosomal control of microtubule dynamics. Proc Natl Acad Sci U.S.A 96: 115–120. doi: 10.1073/pnas.96.1.115

|
| [2] |
Nishita M, Satake T, Minami Y, et al. (2017) Regulatory mechanisms and cellular functions of non-centrosomal microtubules. J Biochem 162: 1–10. doi: 10.1093/jb/mvx018
|
| [3] |
Bartolini F, Gundersen GG (2006) Generation of noncentrosomal microtubule arrays. J Cell Sci 119: 4155–4163. doi: 10.1242/jcs.03227
|
| [4] |
Irina BA, Tatyana B, Gary GB, et al. (2015) Centrosome nucleates numerous ephemeral microtubules and only few of them participate in the radial array. Cell Biol Int 39: 1203–1216. doi: 10.1002/cbin.10492
|
| [5] | Vorobjev IA, Chentsov YS (1983) The dynamics of reconstitution of microtubules around the cell center after cooling. Eur J Cell Biol 30: 149–153. |
| [6] |
Keating TJ, Peloquin JG, Rodionov VI, et al. (1997) Microtubule release from the centrosome. Proc Natl Acad Sci U.S.A 94: 5078–5083. doi: 10.1073/pnas.94.10.5078
|
| [7] |
Waterman-Storer CM, Salmon ED (1997) Actomyosin-based retrograde flow of microtubules in the lamella of migrating epithelial cells influences microtubule dynamic instability and turnover and is associated with microtubule breakage and treadmilling. J Cell Sci 139: 417–434. doi: 10.1083/jcb.139.2.417
|
| [8] | Vorobjev IA, Svitkina TM, Borisy GG (1997) Cytoplasmic assembly of microtubules in cultured cells. J Cell Sci 110: 2635–2645. |
| [9] |
Petry S, Vale RD (2015) Microtubule nucleation at the centrosome and beyond. Nat Cell Biol 17: 1089–1093. doi: 10.1038/ncb3220
|
| [10] |
Vorobjev IA, Alieva IB, Grigoriev IS, et al. (2003) Microtubule dynamics in living cells: Direct analysis in the internal cytoplasm. Cell Biol Int 27: 293–294. doi: 10.1016/S1065-6995(02)00331-1
|
| [11] | Alieva IB, Borisy GG, Vorobjev IA (2008) Spatial organization of centrosome-attached and free microtubules in 3T3 fibroblasts. Cell Tissue Biol 50: 936–946. |
| [12] | Komarova YA, Vorobjev IA, Borisy GG (2002) Life cycle of MTs: Persistent growth in the cell interior, asymmetric transition frequencies and effects of the cell boundary. J Cell Sci 115: 3527–3539. |
| [13] | Vorobjev IA, Rodionov VI, Maly IV, et al. (1999) Contribution of plus and minus end pathways to microtubule turnover. J Cell Sci 112: 2277–2289. |
| [14] |
Komarova YA, Akhmanova AS, Kojima S, et al. (2002) Cytoplasmic linker proteins promote microtubule rescue in vivo. J Cell Biol 159: 589–599. doi: 10.1083/jcb.200208058
|
| [15] | Vorob'Ev IA, Grigor'Ev IS, Borisy GG (2000) Microtubule dynamics in cultured cells. Ontogenez 31: 420–428. |
| [16] |
Matov A, Applegate K, Kumar P, et al. (2010) Analysis of microtubule dynamic instability using a plus-end growth marker. Nat Methods 7: 761–768. doi: 10.1038/nmeth.1493
|
| [17] | Tanaka N, Meng W, Nagae S, et al. (2012) Nezha/CAMSAP3 and CAMSAP2 cooperate in epithelial-specific organization of noncentrosomal microtubules. Proc Natl Acad Sci U.S.A 109: 20029–20034. |
| [18] |
Efimov A, Kharitonov A, Efimova N, et al. (2007) Asymmetric CLASP-dependent nucleation of noncentrosomal microtubules at the trans-Golgi network. Dev Cell 12: 917–930. doi: 10.1016/j.devcel.2007.04.002
|
| [19] | Alieva IB, Zemskov EA, Kireev II, et al. (2010) Microtubules growth rate alteration in human endothelial cells. J Biomed Biotechnol 2010: 671536. |
| [20] |
Salaycik KJ, Fagerstrom CJ, Murthy K, et al. (2005) Quantification of microtubule nucleation, growth and dynamics in wound-edge cells. J Cell Sci 118: 4113–4122. doi: 10.1242/jcs.02531
|
| [21] | Gudima GO, Vorobjev IA, Chentsov YS (1988) Centriolar location during blood cell spreading and motion in vitro: An ultrastructural analysis. J Cell Sci 89: 225–241. |
| [22] |
Alieva IB, Vorobjev IA (1994) Centrosome behavior under the action of a mitochondrial uncoupler and the effect of disruption of cytoskeleton elements on the uncoupler-induced alterations. J Struct Biol 113: 217–224. doi: 10.1006/jsbi.1994.1056
|
| [23] |
Komarova Y, De CG, Grigoriev I, et al. (2009) Mammalian end binding proteins control persistent microtubule growth. J Cell Biol 184: 691–706. doi: 10.1083/jcb.200807179
|
| [24] |
Li W, Moriwaki T, Tani T, et al. (2012) Reconstitution of dynamic microtubules with Drosophila XMAP215, EB1, and Sentin. J Cell Biol 199: 849–862. doi: 10.1083/jcb.201206101
|
| [25] |
Woodruff JB, Gomes BF, Widlund PO, et al. (2017) The Centrosome Is a Selective Condensate that Nucleates Microtubules by Concentrating Tubulin. Cell 169: 1066. doi: 10.1016/j.cell.2017.05.028
|
| [26] |
Wang L, Brown A (2002) Rapid movement of microtubules in axons. Curr Biol 12: 1496–1501. doi: 10.1016/S0960-9822(02)01078-3
|
| [27] |
Doxsey S, Mccollum D, Theurkauf W (2005) Centrosomes in cellular regulation. Annu Rev Cell Dev Biol 21: 411–434. doi: 10.1146/annurev.cellbio.21.122303.120418
|
| [28] |
Rogers GC, Rusan NM, Peifer M, et al. (2008) A multicomponent assembly pathway contributes to the formation of acentrosomal microtubule arrays in interphase Drosophila cells. Mol Biol Cell 19: 3163–3178. doi: 10.1091/mbc.E07-10-1069
|
| [29] |
Luders J, Patel UK, Stearns T (2006) GCP-WD is a gamma-tubulin targeting factor required for centrosomal and chromatin-mediated microtubule nucleation. Nat Cell Biol 8: 137–147. doi: 10.1038/ncb1349
|
| [30] |
Cottam DM, Tucker JB, Rogers-Bald MM, et al. (2006) Non-centrosomal microtubule-organising centres in cold-treated cultured Drosophila cells. Cell Motil Cytoskeleton 63: 88–100. doi: 10.1002/cm.20103
|
| [31] |
Waterman-Storer CM, Worthylake RA, Liu BP, et al. (1999) Microtubule growth activates Rac1 to promote lamellipodial protrusion in fibroblasts. Nat Cell Biol 1: 45–50. doi: 10.1038/9018
|
| [32] |
Rid R, Schiefermeier N, Grigoriev I, et al. (2005) The last but not the least: The origin and significance of trailing adhesions in fibroblastic cells. Cell Motil Cytoskeleton 61: 161–171. doi: 10.1002/cm.20076
|
| [33] |
Prigozhina NL, Waterman-Storer CM (2004) Protein kinase D-mediated anterograde membrane trafficking is required for fibroblast motility. Curr Biol 14: 88–98. doi: 10.1016/j.cub.2004.01.003
|
| [34] |
Shelden E, Wadsworth P (1993) Observation and quantification of individual microtubule behavior in vivo: Microtubule dynamics are cell-type specific. J Cell Biol 120: 935–945. doi: 10.1083/jcb.120.4.935
|
| [35] | Chapin SJ, Bulinski JC (1991) Non-neuronal 210 × 103 Mr microtubule-associated protein (MAP4) contains a domain homologous to the microtubule-binding domains of neuronal MAP2 and tau. J Cell Sci 98: 27–36. |
| [36] |
Kinoshita K, Habermann B, Hyman AA (2002) XMAP215: A key component of the dynamic microtubule cytoskeleton. Trends Cell Biol 12: 267–273. doi: 10.1016/S0962-8924(02)02295-X
|
| [37] |
Toya M, Takeichi M (2016) Organization of Non-centrosomal Microtubules in Epithelial Cells. Cell Struct Funct 41: 127–135. doi: 10.1247/csf.16015
|
| [38] |
Guerin CM, Kramer SG (2009) Cytoskeletal remodeling during myotube assembly and guidance: Coordinating the actin and microtubule networks. Commun Integr Biol 2: 452–457. doi: 10.4161/cib.2.5.9158
|
| [39] |
Oddoux S, Zaal KJ, Tate V, et al. (2013) Microtubules that form the stationary lattice of muscle fibers are dynamic and nucleated at Golgi elements. J Cell Biol 203: 205–213. doi: 10.1083/jcb.201304063
|
| [40] |
Conde C, Caceres A (2009) Microtubule assembly, organization and dynamics in axons and dendrites. Nat Rev Neurosci 10: 319–332. doi: 10.1038/nrn2631
|
| [41] |
Akhmanova A, Steinmetz MO (2015) Control of microtubule organization and dynamics: Two ends in the limelight. Nat Rev Mol Cell Biol 16: 711–726. doi: 10.1038/nrm4084
|
| [42] |
Jiang K, Hua S, Mohan R, et al. (2014) Microtubule minus-end stabilization by polymerization-driven CAMSAP deposition. Dev Cell 28: 295–309. doi: 10.1016/j.devcel.2014.01.001
|
| [43] |
Kamasaki T, O'Toole E, Kita S, et al. (2013) Augmin-dependent microtubule nucleation at microtubule walls in the spindle. J Cell Biol 202: 25–33. doi: 10.1083/jcb.201304031
|
| [44] |
Oakley BR, Paolillo V, Zheng Y (2015) Gamma-Tubulin complexes in microtubule nucleation and beyond. Mol Biol Cell 26: 2957–2962. doi: 10.1091/mbc.E14-11-1514
|
| [45] |
Uehara R, Nozawa RS, Tomioka A, et al. (2009) The augmin complex plays a critical role in spindle microtubule generation for mitotic progression and cytokinesis in human cells. Proc Natl Acad Sci U.S.A 106: 6998–7003. doi: 10.1073/pnas.0901587106
|
| [46] |
Johmura Y, Soung NK, Park JE, et al. (2011) Regulation of microtubule-based microtubule nucleation by mammalian polo-like kinase 1. Proc Natl Acad Sci U.S.A 108: 11446–11451. doi: 10.1073/pnas.1106223108
|
| [47] |
Hsia KC, Wilsonkubalek EM, Dottore A, et al. (2014) Reconstitution of the augmin complex provides insights into its architecture and function. Nat Cell Biol 16: 852–863. doi: 10.1038/ncb3030
|
 genetics-05-02-141-s001.pdf genetics-05-02-141-s001.pdf |
 |
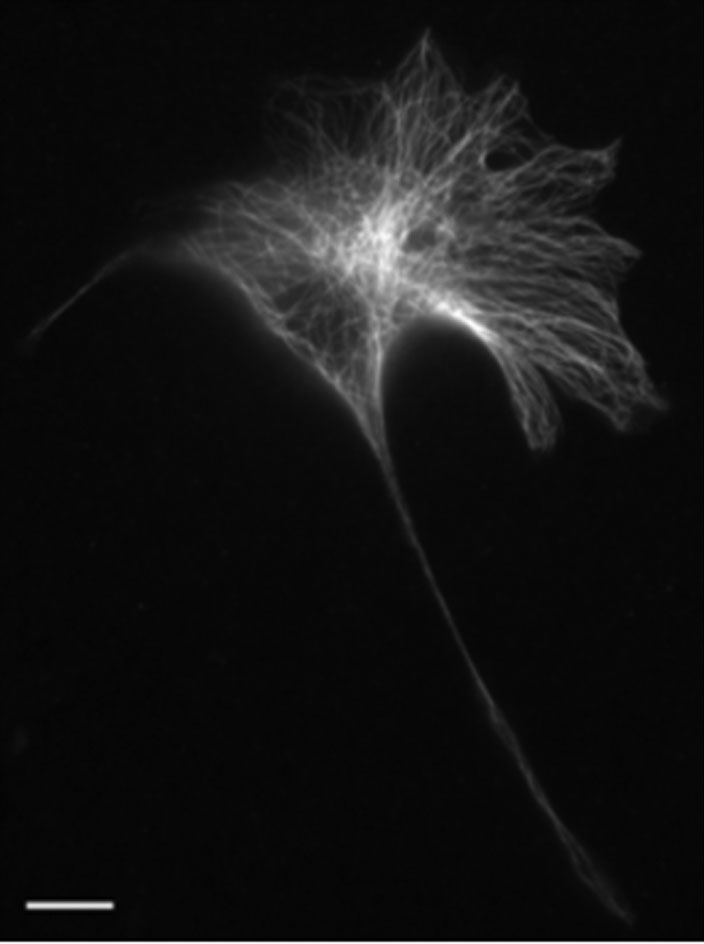
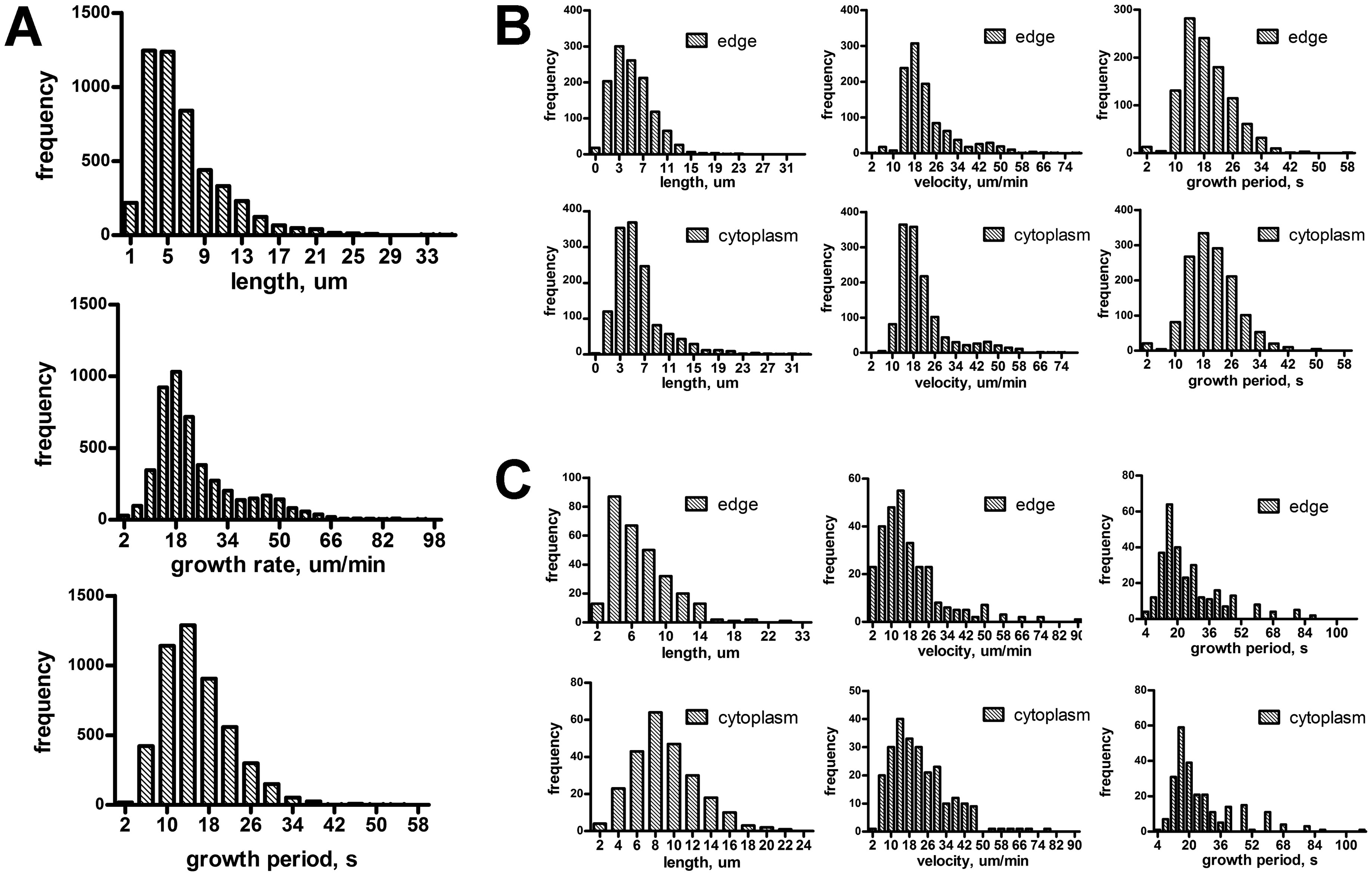
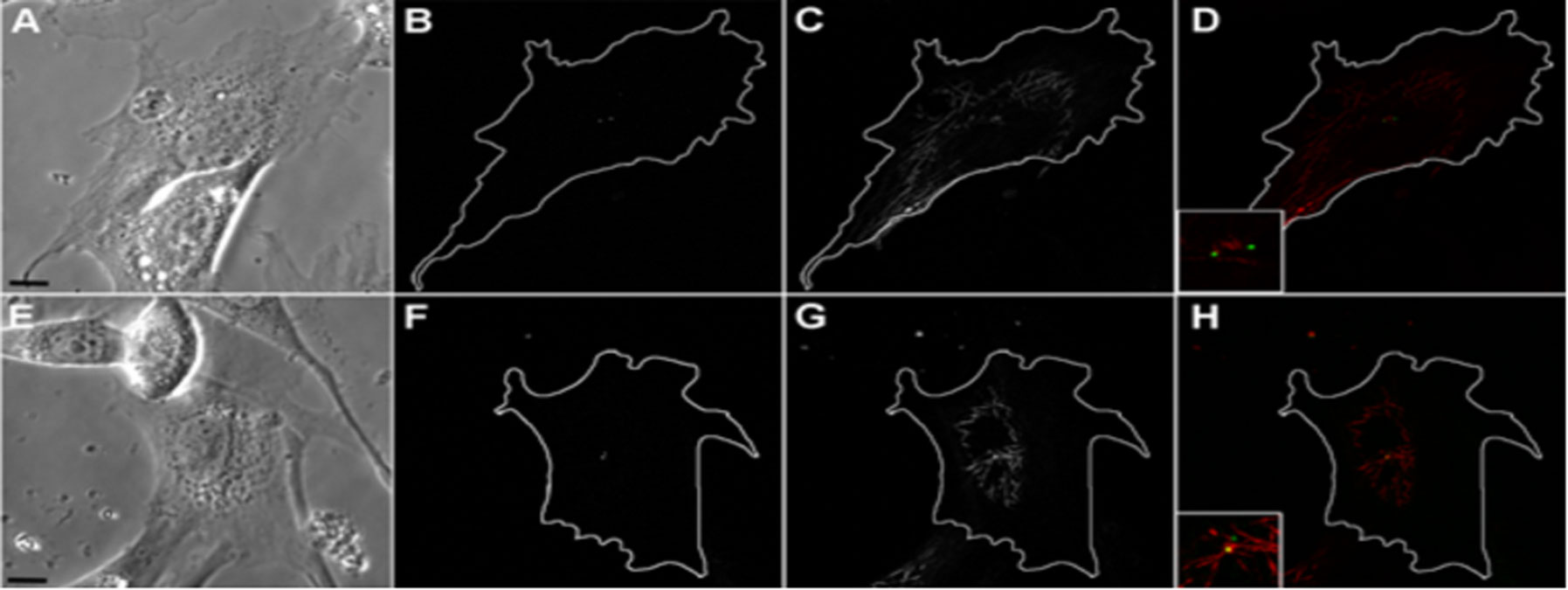
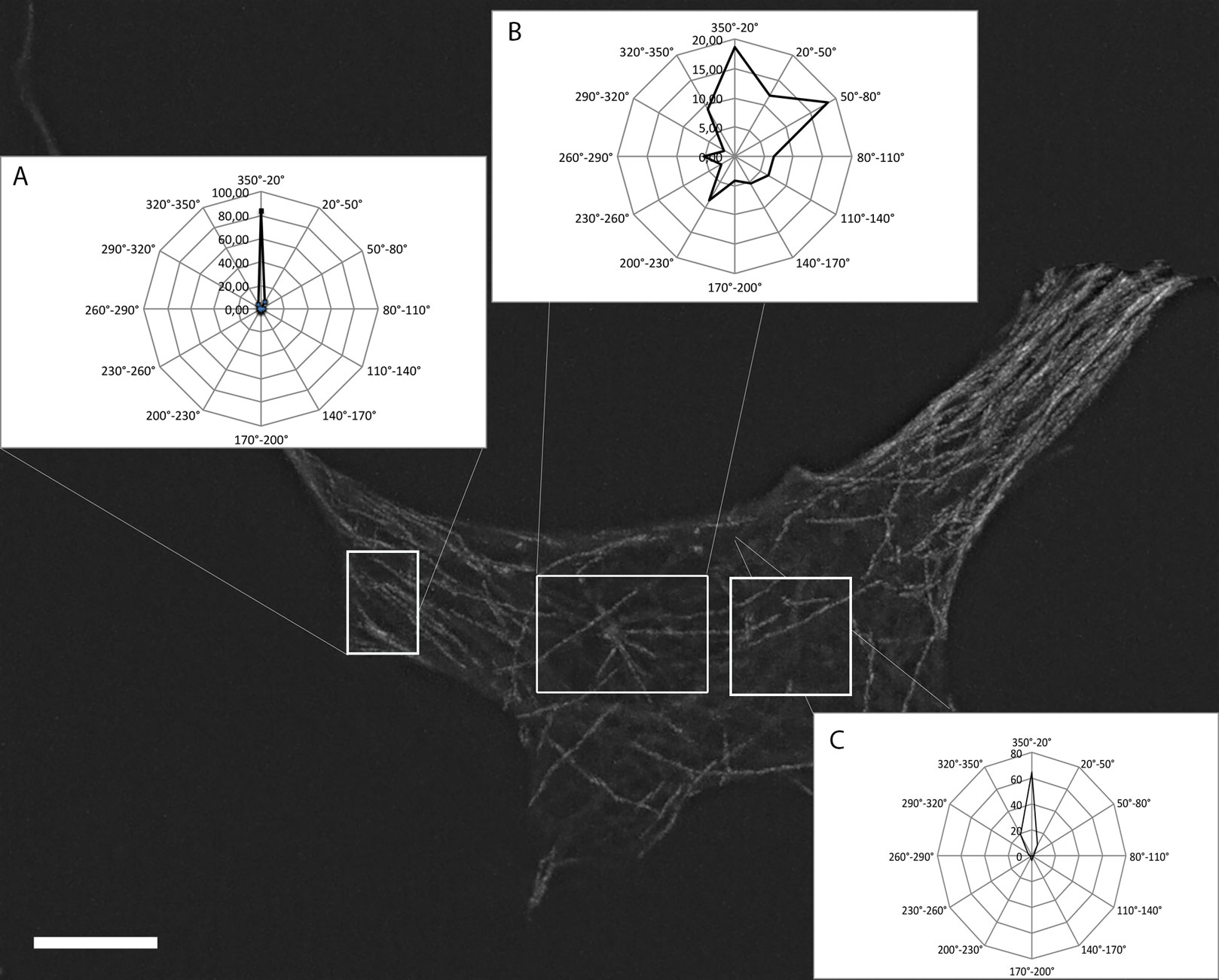
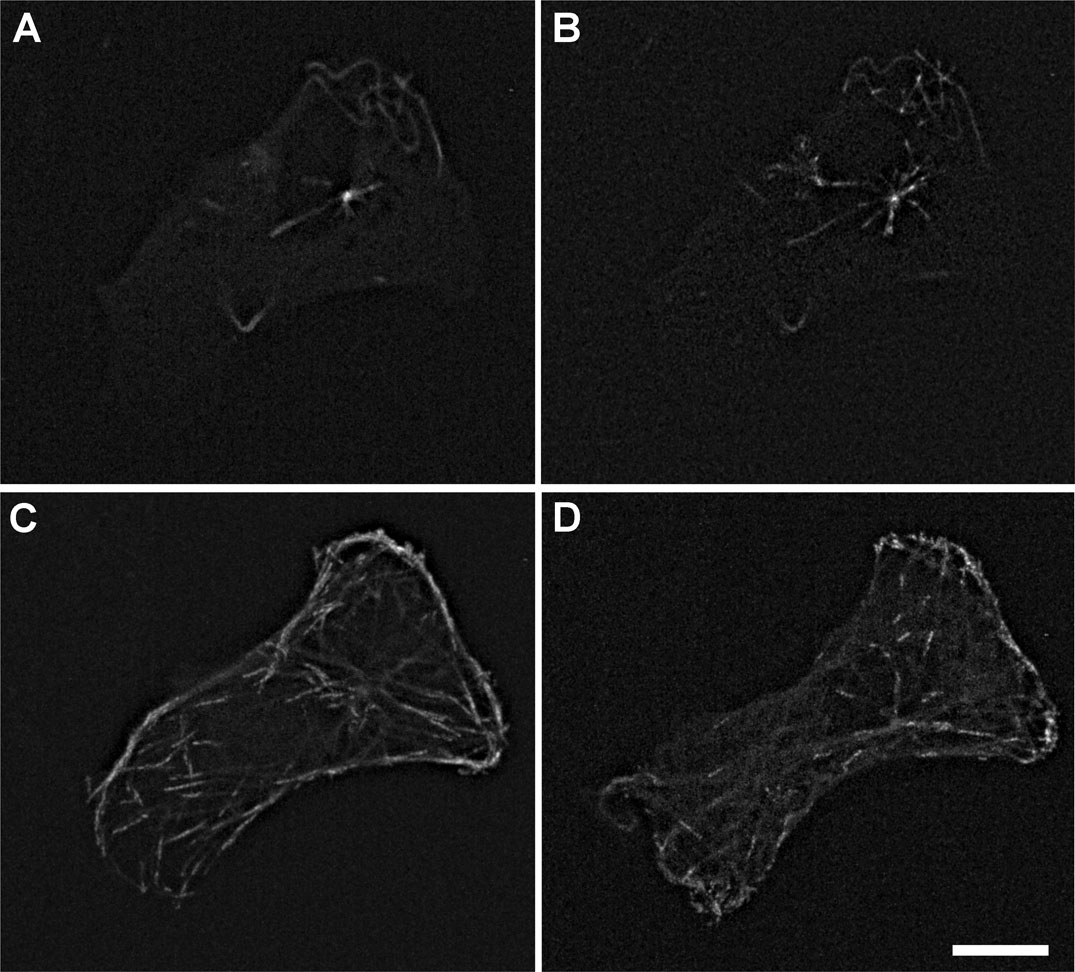
Figures(9) / Tables(1)

Yekaterina Zvorykina, Anna Tvorogova, Aleena Gladkikh, Ivan Vorobjev. Non-centrosomal MTs play a crucial role in organization of MT array in interphase fibroblasts[J]. AIMS Genetics, 2018, 5(2): 141-160. doi: 10.3934/genet.2018.2.141







 DownLoad:
DownLoad: