Citation: Isabella Colombini, Mario Fallaci, Lorenzo Chelazzi. Ecology and trophic links of macroinvertebrates in a dune slack of a Mediterranean coastal ecosystem (Grosseto, Italy)[J]. AIMS Environmental Science, 2017, 4(4): 562-584. doi: 10.3934/environsci.2017.4.562

| [1] | Ranwell D (1960) Newborough Warren, Anglesey: II. Plant associes and succession cycles of the sand dune and dune slack vegetation. J Ecol 48: 117-141. |
| [2] | Ranwell D (1959) Newborough Warren, Anglesey: I. The dune system and dune slack habitat. J. Ecol 47: 571-601. |
| [3] |
Grootjans AP, Ernst WHO, Stuyfzand PJ (1998) European dune slacks: strong interactions of biology, pedogenesis and hydrology. Trends Ecol Evol 13: 96-100. doi: 10.1016/S0169-5347(97)01231-7

|
| [4] |
Lammerts EJ, Grootjans, AP (1997) Nutrient deficiency in dune slack pioneer vegetation: a review. J Coast Conserv 3: 87-94. doi: 10.1007/BF02908183
|
| [5] |
Sival FP, Grootjans AP, Stuyfzand PJ, et al. (1997) Variation in groundwater composition and decalcification depth in a dune slack: effects on basiphilous vegetation. J Coast Conserv 3: 79-86. doi: 10.1007/BF02908182
|
| [6] | Lammerts EJ, Grootjans AP (1998) Key environmental variables determining the occurrence and life span of basiphilous dune slack vegetation. Acta Bot Neerl 47: 369-392. |
| [7] | Etherington JR (1982) Environment and Plant Ecology, 2nd Eds., John Wiley and Sons, Chichester. |
| [8] |
Willis AJ (1963) Braunton Burrows: the effects on the vegetation of the addition of mineral nutrients to the dune soils. J Ecol 51: 353-374. doi: 10.2307/2257690
|
| [9] | Ernst WHO (1991) Ökophysiologie von pflanzen in Küstendünen Europas in einem gradienten von der Nordsee zum Mittelmeer. Ber Reinhold Tüxen Ges 3: 157-172. |
| [10] | Lammerts EJ, Grootjans AP, Stuyfzand PJ, et al. (1995) Endangered dune slack plants; gastronomers in need of mineral water. In: Salman, A.H.P.M., Berends, H., Bonazountas, M. Author, Coastal Management and Habitat Conservation, Vol. I, Leiden: EUCC, 355-369. |
| [11] |
Sival FP (1996) Mesotrophic basiphilous communities affected by changes in soil properties in two dune slack chronosequences. Acta Bot Neerl 45: 95-106. doi: 10.1111/j.1438-8677.1996.tb00498.x
|
| [12] |
Grootjans AP, Engelmoer M, Hendriksma P, et al. (1988) Vegetation dynamics in a wet dune slack I: rare species decline on the Waddenisland of Schiermonnikoog in the Netherlands. Acta Bot Neerl 37: 265-278. doi: 10.1111/j.1438-8677.1988.tb02135.x
|
| [13] | Grootjans AP, Sival FP, Stuyfzand, PJ (1996) Hydro-geochemical analysis of a degraded dune slack. Vegetatio 126: 27-38. |
| [14] | Willis AJ, Folkes BF, Hope-Simpson JF, et al. (1959) Braunton Burrows: the dune system and its vegetation. Part II. J Ecol 47: 249-288. |
| [15] | Martinez ML, Maun MA, Psuty NP (2004). The fragility and conservation of the world's coastal dunes: geomorphological, ecological and socio-economic perspectives. In: Martinez, M.L., Psuty, N.P. Author, Coastal Dunes: Ecology and Conservation. Ecological Studies Berlin: Springer, 355-369. |
| [16] |
Provoost S, Jones MLM, Edmondson SE (2011) Changes in landscape and vegetation of coastal dunes in northwest Europe: a review. J Coast Conserv 15: 207-226. doi: 10.1007/s11852-009-0068-5
|
| [17] | Grootjans AP, Everts H, Bruin K, et al. (2001) Restoration of wet dune slacks on the Dutch Wadden Sea Islands: recolonization after large-scale sod cutting. Restor Ecol 9, 137-146. |
| [18] | Davy AJ, Grootjans AP, Hiscock K, et al. (2006) Development of eco-hydrological guidelines for dune habitats-Phase 1. English Nature Research Reports, No.696, Peterborough. |
| [19] | Davy AJ, Hiscock KM, Jones MLM, et al. (2010) Ecohydrological Guidelines for Wet Dune Habitats-Phase 2. Environ Agency UK Rep |
| [20] | Desenders K, Maelfait J-P, Baert L (2007) Ground beetles as 'early warning-indicators' in restored salt marshes and dune slacks. In: Isermann, M., Kiel, K. Author, Land Use Changes in Europe as a Challenge for Restoration; Ecological, Economical and Ethical Dimensions, 2007. Proceedings of the 5th European Conference on Ecological Restoration. EUCC-The Coastal Union, Greifswald, Germany: 25-39. |
| [21] | van der Hagen HGJM, Geelen LHWT, de Vries CN (2000) Dune slack restoration in Dutch mainland coastal dunes. J Nat Conserv 16:1-11. |
| [22] |
Acosta A, Blasi C, Stanisc A (2000) Spatial connectivity and boundary patterns in coastal dune vegetation in the Circeo National Park, central Italy. J Veg Sci 11: 149-154. doi: 10.2307/3236787
|
| [23] |
Acosta A, Stanisci A, Ercole S, et al. (2003) Sandy coastal landscape of the Lazio region (Central Italy). Phytocoenologia 33: 715-726 doi: 10.1127/0340-269X/2003/0033-0715
|
| [24] |
Carranza ML, Acosta A, Stanisci A, et al. (2008) Ecosystem classification and EU habitat distribution assessment in sandy coastal environments. Environ Monit Assess 140: 99-107. doi: 10.1007/s10661-007-9851-7
|
| [25] |
De Luca E, Novelli C, Barbato F, et al. (2011) Coastal dune systems and disturbance factors: monitoring and analysis in central Italy. Environ Monit Assess 183: 437-450. doi: 10.1007/s10661-011-1931-z
|
| [26] |
Attorre F, Maggini A, Di Traglia M, et al. (2013) A methodological approach for assessing the effects of disturbance factors on the conservation status of Mediterranean coastal dune systems. Appl Veg Sci 16: 333-342. doi: 10.1111/avsc.12002
|
| [27] |
Mazzotti S (1995) Population structure of Emys orbicularis in the Bardello (Po Delta, Northern Italy). Amphibi Reptil 16: 77-85. doi: 10.1163/156853895X00217
|
| [28] | Hammond PM, (2000) Coastal Staphylinidae (rove beetles) in the British Isles, with special reference to saltmarshes. In: Sherwood, B.R., Gardiner, B.G., Harris, T. Author, British Saltmarshes. Cardigan, UK: Forrest Text. 247-302. |
| [29] | Good JA (1999) Intertidal aerobic sandflats as a habitat for marine Coleoptera (Carabidae, Heteroceridae, Staphylinidae) in Ireland. Ir Nat J 26: 73-80. |
| [30] | Eyre MD, Luff ML (2005) The distribution of epigeal beetle (Coleoptera) assemblages on the north-east England Coast. J Coast Res 21: 982-990. |
| [31] |
Colombini I, Brilli M, Fallaci M, et al. (2011) Food webs of a sandy beach macroinvertebrate community using stable isotopes analysis. Acta Oecologica 37: 422-432. doi: 10.1016/j.actao.2011.05.010
|
| [32] |
Hood-Nowotny R, Knols BG (2007) Stable isotope methods in biological and ecological studies of arthropods. Entomol Exp Appl 124:3-16 doi: 10.1111/j.1570-7458.2007.00572.x
|
| [33] | Marques JC, Gonçalves SC, Pardal MA, et al. (2003) Comparison of Talitrus saltator (Amphipoda, Talitridae) biology, dynamics and secondary production in Atlantic (Portugal) and Mediterranean (Italy and Tunisia) populations. Estuar Coast Shelf Sci 58S: 127-148. |
| [34] | McLachlan A, Brown A (2006) The Ecology of Sandy Shores, 2 Eds., London: Academic Press. |
| [35] | Colombini I, Chaouti A, Fallaci M, et al. (2006) Effect of freshwater river discharge on terrestrial arthropods in Atlantic and Mediterranean sandy shores. In: Scapini, F. Author, The Mediterranean Coastal Areas from Watershed to the Sea: interactions and changes. Proceedings of the MEDCORE International Conference, Florence: Florence University Press, 233-261. |
| [36] | Mitsch WJ, Gosselink JG (2007) Wetlands, 4 Eds. John Wiley and Sons, New York. |
| [37] | Vazquez G (2004) The role of algal mats on community succession in dunes and dune slacks. In: Martínez, M.L., Psuty, N.P. Author, Coastal Dunes, Ecology and Conservation. Ecological Studies, vol. 171. Springer-Verlag Berlin Heidelberg, 189-204. |
| [38] | Moreno M, Marin V, Salvo VS, et al (2006) Microbial and meiofaunal communities from a sandy beach in the Maremma Regional Park, Italy. In: Scapini, F. Author, The Mediterranean Coastal Areas from Watershed to the Sea: Interactions and Changes. Proceedings of the MEDCORE International Conference, Florence: Florence University Press, 297-310. |
| [39] |
Colombini I, Fallaci M, Chelazzi L (2015) Ecological strategies of Macarorchestia remyi compared to two sympatric talitrids of a Tyrrhenian beach. Acta Oecol 67: 49-58. doi: 10.1016/j.actao.2015.06.001
|
| [40] |
Mateo MA, Serrano O, Serrano L, et al. (2008) Effects of sample preparation on stable isotope ratios of carbon and nitrogen in marine invertebrates: implications for food web studies using stable isotopes. Oecologia 157: 105-115. doi: 10.1007/s00442-008-1052-8
|
| [41] |
Serrano O, Serrano L, Mateo MA, et al. (2008) Acid washing effect on elemental and isotopic composition of whole beach arthropods: Implications for food web studies using stable isotopes. Acta Oecol 34: 89-96. doi: 10.1016/j.actao.2008.04.002
|
| [42] |
Parnell AC, Inger R, Bearhop S, et al. (2010) Source portioning using stable isotopes: coping with too much variation. PLoS One 5: 9672. doi: 10.1371/journal.pone.0009672
|
| [43] |
Post DM (2002) Using stable isotopes to estimate trophic position: models, methods and assumptions. Ecol 83: 703-718. doi: 10.1890/0012-9658(2002)083[0703:USITET]2.0.CO;2
|
| [44] | ter Braak CJF, Smilauer P (2002) CANOCO reference manual and CanoDraw for Windows user's guide: software for canonical community ordination (version 4.5). Microcomputer Power, Ithaca, New York. |
| [45] | Batschelet E (1981) Circular Statistic in Biology. London: Academic Press. |
| [46] | Davidson AS (2001) Pompilius cinereus (Fabricius, 1775): description and notes. In Bees, Wasps and Ants Recording Society-Trial Dataset. Available from: http://www.bwars.com/wasp/pompilidae/pompilinae/pompilus-cinereus. |
| [47] | McGavin GC (2010) Pocket Nature. Insects and spiders. London: Dorling Kindersley. |
| [48] | Sival FP, Grootjans AP (1996) Dynamics of seasonal bicarbonate supply in a dune slack: effects on organic matter, nitrogen pool and vegetation succession. Veg 126: 39-50 |
| [49] |
Colombini I, Fallaci M, Gagnarli E, et al. (2013) The behavioural ecology of two sympatric talitrid species, Talitrus saltator (Montagu) and Orchestia gammarellus (Pallas) on a Tyrrhenian sandy beach dune system. Estuar Coast Shelf Sci 117: 37-47. doi: 10.1016/j.ecss.2012.09.016
|
| [50] |
Hoback WW, Golick DA, Svatos TM, et al. (2000) Salinity and shade preferences result in ovipositional differences between sympatric tiger beetle species. Ecol Entomol 25: 180-187. doi: 10.1046/j.1365-2311.2000.00256.x
|
| [51] | Pearson DL, Vogler AP (2001) Tiger beetles: the evolution, ecology, and diversity of the cicindelids, New York: Cornell University Press, Ithaca, 333. |
| [52] |
Jaskuła R (2007) Remarks on diversity and distribution of tiger beetles (Coleoptera, Cicindeledae) of Albania. Fragm Faun 50: 127-138. doi: 10.3161/00159301FF2007.50.2.127
|
| [53] |
Jaskuła R (2011) How unique is the tiger beetle fauna (Coleoptera, Cicindelidae) of the Balkan Peninsula? ZooKeys 100: 487-502. doi: 10.3897/zookeys.100.1542
|
| [54] |
Jaskuła R (2015) The Maghreb-one more important biodiversity hot spot for tiger beetle fauna (Coleoptera, Carabidae, Cicindelinae) in the Mediterranean region. ZooKeys 482: 35-53. doi: 10.3897/zookeys.482.8831
|
| [55] |
Howe MA, Knight GT, Clee C (2010) The importance of coastal sand dunes for terrestrial invertebrates in Wales and the UK, with particular reference to aculeate Hymenoptera (bees, wasps and ants). J Coast Conserv 14: 91-102. doi: 10.1007/s11852-009-0055-x
|
| [56] |
Rodríguez-Flores PC, Gutiérrez-Rodríguez J, Aguirre-Ruiz E, et al. (2016) Salt lakes of La Mancha (Central Spain): A hot spot for tiger beetle (Carabidae, Cicindelinae) species diversity. ZooKeys 561: 63-103. doi: 10.3897/zookeys.561.6042
|
| [57] |
Schultz TD (1989) Habitat preferences and seasonal abundances of eight sympatric species of tiger beetle, genus Cicindela (Coleoptera: Cicindelidae), in Bastrop State Park, Texas. Southwest Nat 34: 468-477. doi: 10.2307/3671504
|
| [58] |
Hoback WW, Golick DA, Svatos TM, et al. (2000) Salinity and shade preferences result in ovipositional differences between sympatric tiger beetle species. Ecol Entomol 25: 180-187. doi: 10.1046/j.1365-2311.2000.00256.x
|
| [59] |
Satoh A, Hori M (2005) Microhabitat segregation in larvae of six species of coastal tiger beetles in Japan. Ecol Res 20: 143-149. doi: 10.1007/s11284-004-0028-3
|
| [60] | Marcuzzi G (2012) European ecosystems, Junk Publishers, The Hague-Boston-London. |
| [61] |
Fallaci M, Colombini I, Palesse L, et al. (1997) Spatial and temporal strategies in relation to environmental constraints of four tenebrionids inhabiting a Mediterranean coastal dune system. J Arid Environ 37: 45-64. doi: 10.1006/jare.1997.0258
|
| [62] | Framenau V, Reich M, Plachter H (1996) Zum wanderverhalten und zur nahrungso¨kologie von Arctosa cinerea (Fabricius, 1777) (Araneae: Lycosidae) in einer alpinen wildflusslandschaft. Verh Ges Ökol 26: 369-376. |
| [63] | Framenau V, Dieterich M, Reich M, et al. (1996) Life cycle, habitat selection and home ranges of Arctosa cinerea (Fabricius, 1777) (Araneae: Lycosidae) in a braided section of the Upper Isar (Germany, Bavaria). Rev Suisse Zool hors serie: 223-234. |
| [64] | Day MC (1981) A revision of Pompilus (Fabricius) (Hymenoptera: Pompilidae), with further nomenclatural and biological considerations. Bull Br Mus Nat Hist 42: 1-41. |
| [65] | Conti E, Costa G, Petralia A, et al. (2012) Eco-ethology of Parallelomorphus laevigatus (Coleoptera, Carabidae): a species to protect. Atti Fauna Sicil 11: 41-49. |
| [66] |
Bluthgen N, Gebauer G, Fiedler K (2003) Disentangling a rainforest food web using stable isotopes: dietary diversity in a species-rich ant community. Oecologia 137:426-435. doi: 10.1007/s00442-003-1347-8
|
| [67] | Tuzun A (2001) Ecological studies on Vespidae, Eumenidae, Sphecidae, Scoliidae and Apidae species (Insecta: Hymenoptera) from various localities of Turkey. F.U. Fen ve Muh Bilimleri Dergisi 13: 39-47. |
| [68] | Vereecken N, Carriere J (2003) Contribution à l'étude éthologique de la grande Scolie à front jaune Megascolia maculata flavifrons (F.,1775) (Hymenoptera, Scoliidae) en France méditerranéenne. Notes Fauniques Gembloux 53: 71-80. |
| [69] | Barratt, BIP (2003) Aspects of reproductive biology and behaviour of scoliid wasps. DoC Sci Intern Ser 147. |
| [70] | Generani M, Pagliano G, Scaramozzino PL (2006) Hymenoptera Scolioidea (Tiphiidae excluded). In: Ruffo, S. Stoch, F. Author, Checklist and Distribution of the Italian Fauna. 10000 Terrestrial and Inland Water Species. Memorie del Museo Civico di Storia Naturale di Verona. Sezione Scienze della Vita 17: 273-274. |
| [71] | Grandi G (1961) Studi di un entomologo sugli imenotteri superiori. Boll Ist Entomol Univ Bologna 25: 1-659. |
| [72] | Tormos J, Asís JD, Gayubo SF (2003) Description of the mature larva of Nemka viduata (Pallas) (Hymenoptera: Mutillidae: Mutillinae), a parasitoid of Stizus continuus (Klug)(Hymenoptera: Crabronidae: Bembicinae). J Entomol Sci 38: 502-510. |
| [73] |
Brothers DJ, Tschuch G, Burger F (2000) Associations of mutillid wasps (Hymenoptera, Mutillidae) with eusocial insects. Insectes Soc 47: 201-211. doi: 10.1007/PL00001704
|
| [74] | Brothers DJ, (1989) Alternative life-history styles of mutillid wasps (Insecta, Hymenoptera), In: Bruton, M.N. Author, Alternative Life-history Styles of Animals, Dordrecht: Kluwer Academic Publishers, 279-291. |
| [75] |
Polidori C, Mendiola P, Asís JD, et al. (2010) Temporal asynchrony and spatial co-occurrence with the host: the foraging patterns of Nemka viduata, a parasitoid of digger wasps (Hymenoptera: Mutillidae and Crabronidae). J Ethol 28: 353-361. doi: 10.1007/s10164-009-0196-2
|
| [76] |
Scapini F, Chelazzi L, Colombini I, et al. (1992) Surface activity, zonation and migrations of Talitrus saltator (Montagu, 1808) on a Mediterranean beach. Mar Biol 112: 573-581. doi: 10.1007/BF00346175
|
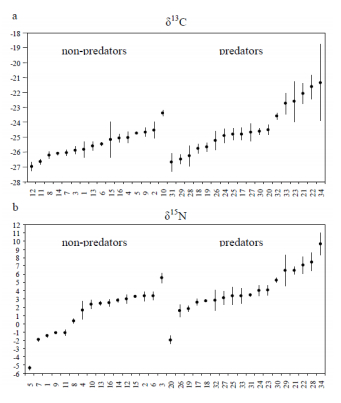
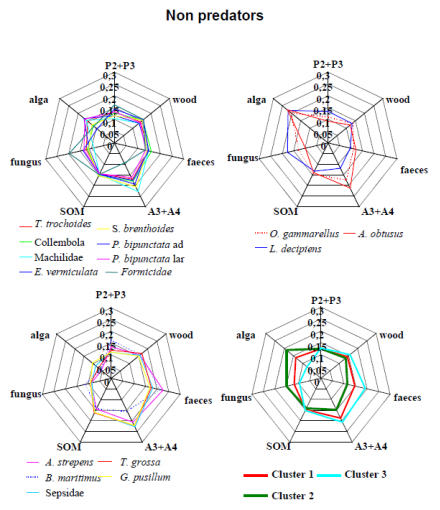
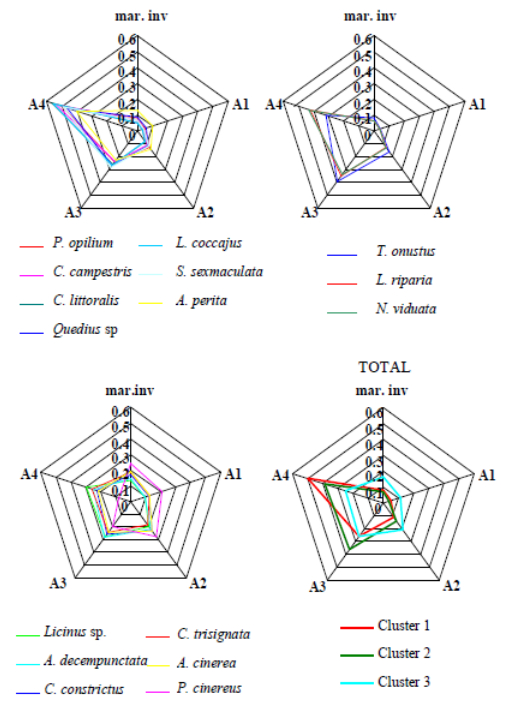
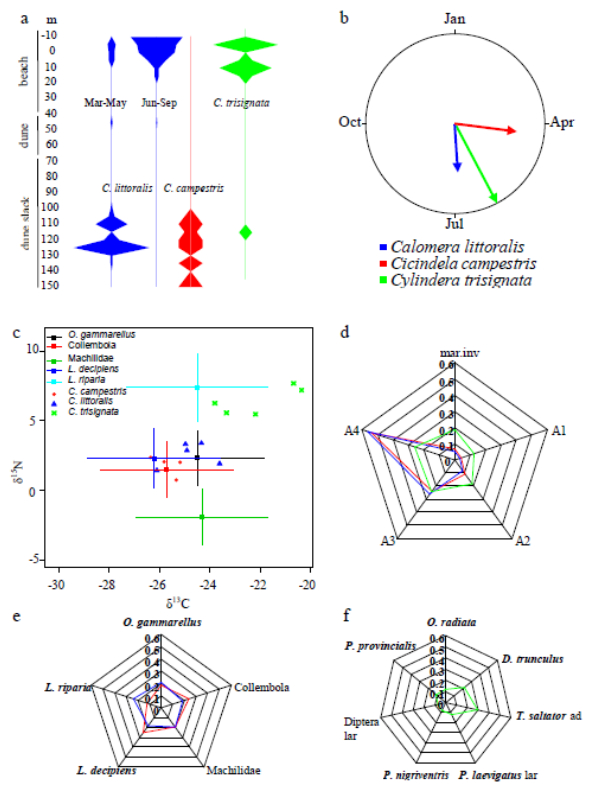
Figures(11) / Tables(1)

Isabella Colombini, Mario Fallaci, Lorenzo Chelazzi. Ecology and trophic links of macroinvertebrates in a dune slack of a Mediterranean coastal ecosystem (Grosseto, Italy)[J]. AIMS Environmental Science, 2017, 4(4): 562-584. doi: 10.3934/environsci.2017.4.562







 DownLoad:
DownLoad: