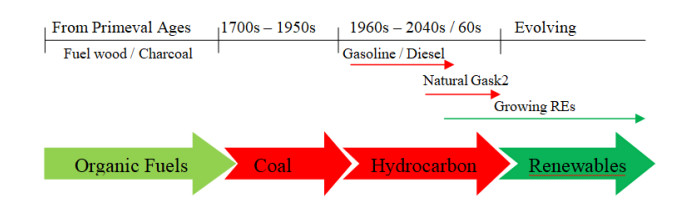
The next transformation in human civilization is occurring in the energy sector. The transition is supported mostly by concern for environmental safety (climate change). Other factors such as energy security and economics also dictate the shift. The transition will be more disruptive to oil-exporting economies such as Nigeria, which depend heavily on it for economic growth. Historical inferences from past energy transitions reveals that discovery and massive acceleration of new energy resources alters mode of production, energy consumption, economic innovation and unlocks multiplier effects on the society. Technology and availability of energy resources is the critical enabler of past transitions. Renewable energy technologies are becoming more cost-competitive relative to fossil fuels. The increasing transition will have serious socioeconomic and political implications, if the country does not diversify its economy. For example, contraction in oil export because of global transition to renewable energy will reduce Nigeria government's revenue, increase unemployment, decrease capacity to finance infrastructure and development projects, and increase poverty rate. Nigeria has enormous renewable energy potential—solar, wind, biomass, etc. The use of renewables will improve environmental quality. For example, increased use of biomass (organic wastes) for energy production will advance Nigeria solid waste management system and vice versa. All these are associated with sustainable resource management.
Citation: Chukwuebuka Okafor, Christian Madu, Charles Ajaero, Juliet Ibekwe, Happy Bebenimibo, Chinelo Nzekwe. Moving beyond fossil fuel in an oil-exporting and emerging economy: Paradigm shift[J]. AIMS Energy, 2021, 9(2): 379-413. doi: 10.3934/energy.2021020
The next transformation in human civilization is occurring in the energy sector. The transition is supported mostly by concern for environmental safety (climate change). Other factors such as energy security and economics also dictate the shift. The transition will be more disruptive to oil-exporting economies such as Nigeria, which depend heavily on it for economic growth. Historical inferences from past energy transitions reveals that discovery and massive acceleration of new energy resources alters mode of production, energy consumption, economic innovation and unlocks multiplier effects on the society. Technology and availability of energy resources is the critical enabler of past transitions. Renewable energy technologies are becoming more cost-competitive relative to fossil fuels. The increasing transition will have serious socioeconomic and political implications, if the country does not diversify its economy. For example, contraction in oil export because of global transition to renewable energy will reduce Nigeria government's revenue, increase unemployment, decrease capacity to finance infrastructure and development projects, and increase poverty rate. Nigeria has enormous renewable energy potential—solar, wind, biomass, etc. The use of renewables will improve environmental quality. For example, increased use of biomass (organic wastes) for energy production will advance Nigeria solid waste management system and vice versa. All these are associated with sustainable resource management.

| [1] |
Falcone PM, Lopolito A, Sica E (2017) Policy mixes towards sustainability transition in the Italian biofuel sector: Dealing with alternative crisis scenarios. Energy Res Soc Sci 33: 105-114. doi: 10.1016/j.erss.2017.09.007

|
| [2] |
Falcone PM (2018) Analysing stakeholders' perspectives towards a socio-technical change: The energy transition journey in Gela municipality. AIMS Energy 6: 645-657. doi: 10.3934/energy.2018.4.645
|
| [3] |
Kokkinos K, Karayannis V, Moustakas K (2020) Circular bio-economy via energy transition supported by Fuzzy Cognitive Map modeling towards sustainable low-carbon environment. Sci Total Environ 721: 1-16. doi: 10.1016/j.scitotenv.2020.137754
|
| [4] |
Okafor C, Ajaero C, Madu C, et al. (2020) Implementation of circular economy principles in management of end-of-life tyres in a developing country (Nigeria). AIMS Environ Sci 7: 406-433. doi: 10.3934/environsci.2020027
|
| [5] |
Thombs RP (2019) When democracy meets energy transitions: A typology of social power and energy system scale. Energy Res Soc Sci 52: 159-168. doi: 10.1016/j.erss.2019.02.020
|
| [6] | Owen R, Brennan G, Lyon F (2018) Enabling investment for the transition to a low carbon economy: Government policy to finance early stage green innovation. Submitted to Current Opinion in Environmental Sustainability Special Issue for the Intergovernmental Panel on Climate Change (IPCC) Sixth Assessment Cycle and Special Report on Global Warming. |
| [7] | Kah M (2018) Electric vehicles and their impact on oil demand: Why forecasts differ. Center on Global Energy Policy, Columbia SIPA, 1-15. |
| [8] | Stevens P (2019) The geopolitical implications of future oil demand, Research Paper. The Royal Institute of International Affairs: Chatham House, 1-42. |
| [9] | Center for Sustainable Systems, University of Michigan (2016) Greenhouse Gases Factsheet. Pub. No. CSS05-21. Available from: https://css.umich.edu/factsheets/greenhouse-gases-factsheet |
| [10] | IPCC (2013) Summary for policymakers. In: Stocker TF, Qin D, Plattner GK, et al. (Eds.), In: Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA. |
| [11] | Johnston RJ, Blakemore R, Bell R (2020) The role of oil and gas companies in the energy transition. Atlantic Council, Washington DC: USA, 1-44. |
| [12] |
Maji IK (2015) Does clean energy contribute to economic growth? Evidence from Nigeria. Energy Rep 1: 145-150. doi: 10.1016/j.egyr.2015.06.001
|
| [13] |
Adewuyi OB, Kiptoo MK, Afolayan AF, et al. (2020) Challenges and prospects of Nigeria's sustainable energy transition. Energy Rep 6: 993-1009. doi: 10.1016/j.egyr.2020.04.022
|
| [14] |
Fouquet R (2016) Historical energy transitions: speed, prices and system transformation. Energy Res Soc Sci 22: 7-12. doi: 10.1016/j.erss.2016.08.014
|
| [15] | Oxford Institute for Energy Studies. The rise of renewables and energy transition: what adaptation strategy for oil companies and oil-exporting countries? (2018) OIES Paper: MEP 19. 1-25. Available from: https://www.oxfordenergy.org/publications/rise-renewables-energy-transition-adaptation-strategy-oil-companies-oil-exporting-countries/. |
| [16] | Pistelli L (2020) Addressing Africa's energy dilemma. In: Hafner and Tagliapietra (eds), The geopolitics of the global energy transition, Lecture Notes in Energy 73: 151-174. |
| [17] | Sovacool BK (2016) The history and politics of energy transitions: Comparing contested views and finding common ground. United Nations University World Institute for Development Economics Research, WIDER working paper 2016/81, Helsinki, Finland. |
| [18] |
Zou C, Zhao Q, Zhang G, et al. (2016) Energy revolution: From a fossil fuel energy era to a new energy era. Nat Gas Ind B 3: 1-11. doi: 10.1016/j.ngib.2016.02.001
|
| [19] | Unger RW (2013) Energy transitions in history: Global cases of continuity and change. Rachel Carson Center Perspectives, 2013/2. |
| [20] | Hafner M, Tagliapietra S (2020) The geopolitics of the global energy transition. In: Lecture Notes in Energy 73: 1-398. |
| [21] | Bressand A (2012) The changed geopolitics of energy and climate and the challenge for Europe. A geopolitical and European perspective on the triple agenda of competition, energy security and sustainability. In: Clingendael international energy programme (CIEP), CIEP paper 04. |
| [22] | REN21, Renewable Energy Policy Network for the 21st Century (2019) Renewables 2019 Global Status Report (Paris: REN21 Secretariat). ISBN 978-3-9818911-7-1. 1-336. |
| [23] | IRENA (2018) Global energy transformation: A roadmap to 2050. International Renewable Energy Agency, Abu Dhabi. 1-76. ISBN 978-92-9260-059-4. |
| [24] | Todd J, Thorstensen L (2013) Creating the clean energy economy: Analysis of the electric vehicle industry. International Energy Development Council, Washington DC: USA, 1-100. |
| [25] | Global EV Outlook 2013, Global EV outlook: Understanding the electric vehicle landscape to 2020. Clean Energy Ministerial, Electric Vehicles Initiative and International Energy Agency. |
| [26] | BP (2020) Statistical review of world energy 2020, 69th edition. Available from: https://www.bp.com/content/dam/bp/business-sites/en/global/corporate/pdfs/energy-economics/statistical-review/bp-stats-review-2020-full-report.pdf. |
| [27] | IRENA, International Renewable Energy Agency (2016) Renewable energy benefits: Measuring the economics, Abu Dhabi: UAE, 1-92. |
| [28] |
Endomah N, Foulds C, Jones A (2016) Energy transitions in Nigeria: The evolution of energy infrastructure provision (1800-2015). Energies 9: 484. doi: 10.3390/en9070484
|
| [29] | Porter G (2012) Reflections on a century of road transport developments in West Africa and their (gendered) impacts on the rural poor. EchoGeo 20: 1-16. |
| [30] | Abioye O, Shubber K, Koenigsberger J (2016) Evaluating the role and impact of railway transport in the Nigerian economy, options and choices: Case of Nigerian Railway Corporation. AshEse J Econ 2: 103-113. |
| [31] | Udosen C, Etok A-I, George IN (2009) Fifty years of oil exploration in Nigeria: The paradox of plenty. Glob J Soc Sci 8: 37-47. |
| [32] | World Bank (2017) Nigeria federal roads development project (P090135) Report No: ICR00004170. |
| [33] | EPA (2010) Landfill recovery and use in Nigeria (Pre-feasibility studies of using LFGE). Grant Number: XA83367801, US Environmental Protection Agency and Centre for People and Environment (CPE). |
| [34] | US Energy Information Administration, Biomass explained: Waste-to-energy (Municipal solid waste), 2019. Available from: https://www.eia.gov/energyexplained/biomass/waste-to-energy.php. |
| [35] | Clean technology investment plan for Nigeria: Updated Note, July 2014, 1-36. |
| [36] | D'Adamo I, Falcone PM, Gastaldi M, et al. (2020) The economic viability of photovoltaic systems in public buildings: Evidence from Italy. Energy 207: 1-10. |
| [37] | PwC, PriceWaterCoopers (2016), Nigeria: Looking beyond oil, 1-32. |
| [38] | Raheem I (2016) Analysis of the effects of oil and non-oil export on economic growth in Nigeria. Hal-01401103v2. |
| [39] | World Bank (2017) Nigeria Bi-annual Economic update: Fragile economy. The World Bank IBRD-IDA No. 1- April 2017. |
| [40] | Oyekanmi S (2020) Updated: Nigerian economy grows by 2.27% in 2019, post highest quarterly growth since 2016 recession. Available from: https://nairametrics.com/2020/02/24/nigerian-economy-grows-by-2-27-post-highest-quarterly-growth-since-2016-recession/. |
| [41] | World Bank (2020) Nigeria's economy faces worst recession in four decades, says new World Bank Report. Available from: https://www.worldbank.org/en/news/press-release/2020/06/25/nigerias-economy-faces-worst-recession-in-four-decades-says-new-world-bank-report. |
| [42] | Auty RM (2001) Introduction and overview. In Auty RS (Ed.), Resource abundance and economic development (1st ed.), Oxford: Oxford University Press, 3-16. |
| [43] | Rosser A (2006) The political economy of the resource curse: A literature survey. IDS Working Paper 268. |
| [44] |
Roche MY, Verolme H, Agbaegbu C, et al. (2020) Achieving sustainable development goals in Nigeria's power sector: Assessment of transition pathways. Clim Policy 20: 846-865. doi: 10.1080/14693062.2019.1661818
|
| [45] | U.S. EIA, Energy Information Administration, (2020) Country analysis executive summary: Nigeria. 1-10. |
| [46] | Okanlawon L (2015) The potential of Nigeria's residential solar rooftop systems. Available from: https://www.renewableenergyworld.com/storage/the-potential-of-nigerias-residential-solar-rooftop-systems/. |
| [47] | Ministry of Mines and Steel Development, nd, Coal. Available from: https://www.minesandsteel.gov.ng/portfolio/coal/. |
| [48] | Chukwu M, Folayan CO, Pam GY, et al. (2016) Characterization of some Nigerian coals for power generation. J Combust, 2016. |
| [49] |
Ogunsola OI (1991) Coal production and utilization trends in Nigeria. Fuel Sci Tech Int 9: 1211-1222. doi: 10.1080/08843759108942323
|
| [50] | Index Mundi, nd, Nigeria coal production and consumption by year (Thousand short tons). Available from: https://www.indexmundi.com/energy/?country=ng&product=coal&graph=production+consumption. |
| [51] | MGI, McKinsey Global Institute (2014) Nigeria's renewal: Delivering inclusive growth in Africa's largest economy. |
| [52] | Dorian JP, Shealy MT, Simberk DR (2020) The global energy transition: Where do we go from here? IAEE Energy Forum/Second Quarter 2020: 11-18. |
| [53] | World Bank, Nigeria's flaring reduction target: 2020 (2017). The World Bank IBRD-IDA. Available from: https://www.worldbank.org/en/news/feature/2017/03/10/nigerias-flaring-reduction-target. |
| [54] | Index mundi (2014) Nigeria crude oil production and consumption by year (Thousand barrels per day). Available from: https://www.indexmundi.com/energy/?country=ng&product=oil&graph=production+consumption. |
| [55] | Statista (2020) Oil production in Nigeria from 1998 to 2019. Available from: https://www.statista.com/statistics/265195/oil-production-in-nigeria-barrels-per-day/. |
| [56] | TheGlobalEconomy.com (2020) Nigeria foreign direct investment, percent of GDP, chart. Available from: https://www.theglobaleconomy.com/Nigeria/Foreign_Direct_Investment/. |
| [57] | UNCTAD, United Nations Conference on Trade and Development, (1999) World investment report 1999. United Nations, New York and Geneva. Available from: http://unct.ad.org/en/docs/wir1999_en.pdf. |
| [58] |
Kareem SD, Kari F, Alam GM, et al. (2012) Foreign direct investment into oil sector and economic growth in Nigeria. Int J Appl Econ Finance 6: 127-135. doi: 10.3923/ijaef.2012.127.135
|
| [59] | Nordea (2020) Foreign direct investment (FDI) in Nigeria. Available from: https://www.nordeatrade.com/en/explore-new-market/nigeria/investment. |
| [60] | Scholten D (ed) (2018) The geopolitics of renewables. Springer, New York. |
| [61] | Spiegel (2018) Neuzulassungen: Erstmalsmehralseine Million E-Autos (New Registrations: First time more than one Million Electric Vehicles). Available from: http://www.spi'egel.de/auto/aktuell/emobilitaet-erstmals-ueber-eine-million-e-autoszugelassen-a-1197636.html. |
| [62] | Couture TD, Leidreiter A (2014) How to achieve 100% renewable energy. World Future Council. Mexikoring 29, 22997 Hamburg, Germany, 1-60. |
| [63] | Seplat Petroleum Development Company (2019) Nigeria's energy transformation: Special feature, Seplat Petroleum Development Company Plc Annual Report and Accounts 2019, 44-51. |
| [64] | World Bank, Ecofys and Vivid Economics (2017) State and Trends of Carbon Pricing 2017 (November), by World Bank, Washington, DC. Doi: 10.1596/978-1-4648-1218-7License:CreativeCommonsAttributionCCBY3.0IGO. |
| [65] | World Economic Forum (2014) Global energy architecture performance index: The new energy architecture challenge—Balancing the energy triangle. |
| [66] | Ministry of Power, Federal Republic of Nigeria (2015) National Renewable Energy and Efficiency Policy (NREEEP) approved by FEC for the electricity sector. |
| [67] | Falobi EO (2020) The role of renewable in Nigeria's energy policy mix. Int Assoc Energy Econ (IAEE) Energy Forum/First Quarter 2020: 41-46. |
| [68] | Offgrid Nigeria (2020) CBN aims for $10m savings on import substitution for solar equipment. Available from: https://www.offgridnigeria.com/cbn-10m-savings-import-substitution-solar-equipment/. |
| [69] |
Asumadu-Sarkodie S, Owusu PA (2016) A review of Ghana's solar energy potential. AIMS Energy 4: 675-696. doi: 10.3934/energy.2016.5.675
|
| [70] |
Akuru UB (2017) Towards 100% renewable energy in Nigeria. Renew Sustain Energy Rev 71: 943-953. doi: 10.1016/j.rser.2016.12.123
|
| [71] |
Esae OE, Sarah J, Mofe A (2020) A critical analysis of the role of energy generation from municipal solid waste (MSW). AIMS Environ Sci 7: 387-405. doi: 10.3934/environsci.2020026
|
| [72] | Bala-Gbogbo E (2012) Nigeria sovereign wealth fund to start investing in March. Available from: https://www.bloomberg.com/news/articles/2012-12-20/nigeria-sovereign-wealth-fund-to-start-investing-in-march. |
| [73] | NSIA, Nigeria Sovereign Investment Authority, (2016) Available from: https://nsia.com.ng/investments. |
| [74] | Eluhaiwe PN (2013) Government role in promoting renewable energy, Central Bank of Nigeria, 1-21. |
| [75] | PwC (2019) X-raying the Nigerian palm oil sector, 1-8. Available from: https://www.pwc.com/ng/en/publications/x-raying-the-nigerian-palm-oil-sector.html. |
| [76] | PwC (2016) Powering Nigeria for the future: The power sector in Nigeria July 2016. |
| [77] | GBI (2013) Nigeria to spend $950 million on generator imports by 2020, viewed 10 July 2018. |
| [78] | The Nation Online News, Nigeria is highest generator importer in Africa, 27 August, 2014. Available from: https://thenationonlineng.net//nigeria-is-highest-generator-importer-in-africa/. |
| [79] | Central Bank of Nigeria (CBN) (2015) Analysis of energy market conditions in Nigeria' Occasional Paper No. 55. |
| [80] | Greenstone M, Reguant M, Ryan N, et al. (2019) Energy and environment: Evidence paper. International Growth Center (IGC), London School of Economic and Political Science, Houghton Street, London WC2A 2AE, 1-44. |
| [81] | Obafemi FN, Ifere EO (2013) Non-technical losses, energy efficiency and conservative methodology in the electricity sector of Nigeria: The case of Calabar, Cross River State. Int J Energy Econ Policy 3: 185-192. |
| [82] | Drücke O (2018) Demand-Side Survey and Analysis of Mid-Sized Power Consumers in Nigeria. Let's Make Solar Work, A Nigerian-German Initiative: Lagos, Nigeria. |




Figures(5) / Tables(3)

Chukwuebuka Okafor, Christian Madu, Charles Ajaero, Juliet Ibekwe, Happy Bebenimibo, Chinelo Nzekwe. Moving beyond fossil fuel in an oil-exporting and emerging economy: Paradigm shift[J]. AIMS Energy, 2021, 9(2): 379-413. doi: 10.3934/energy.2021020







 DownLoad:
DownLoad: