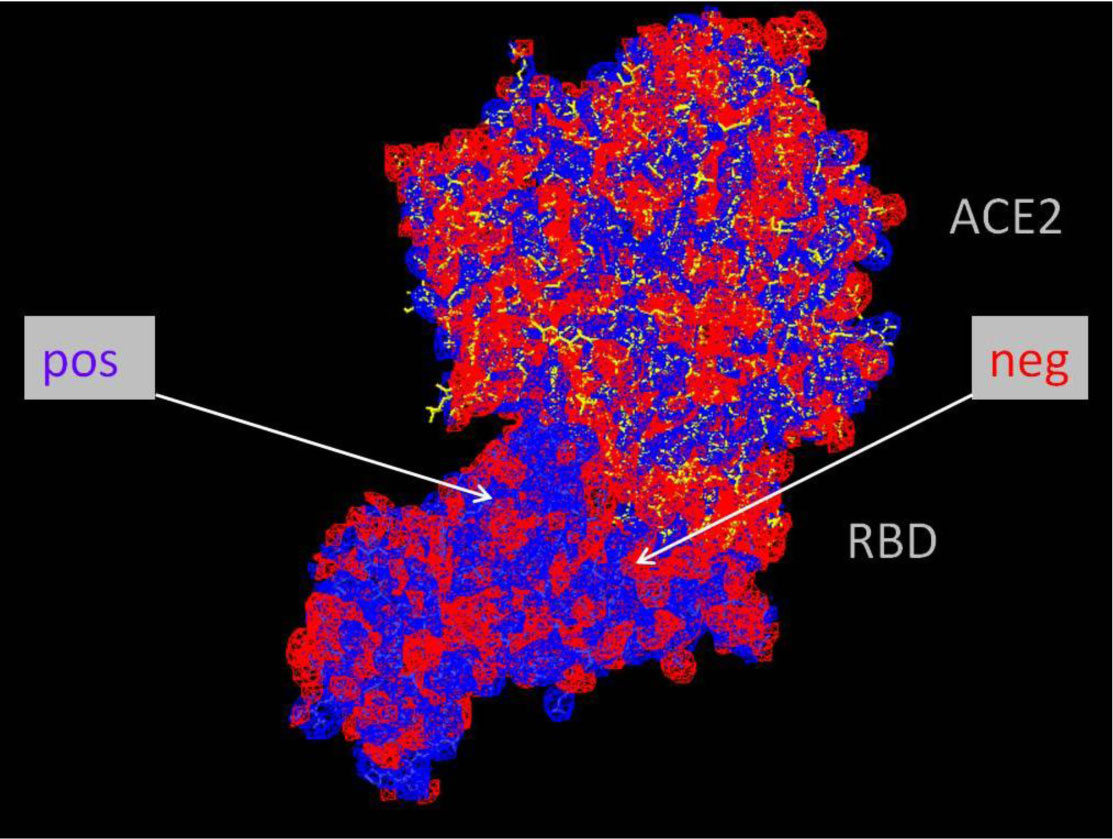
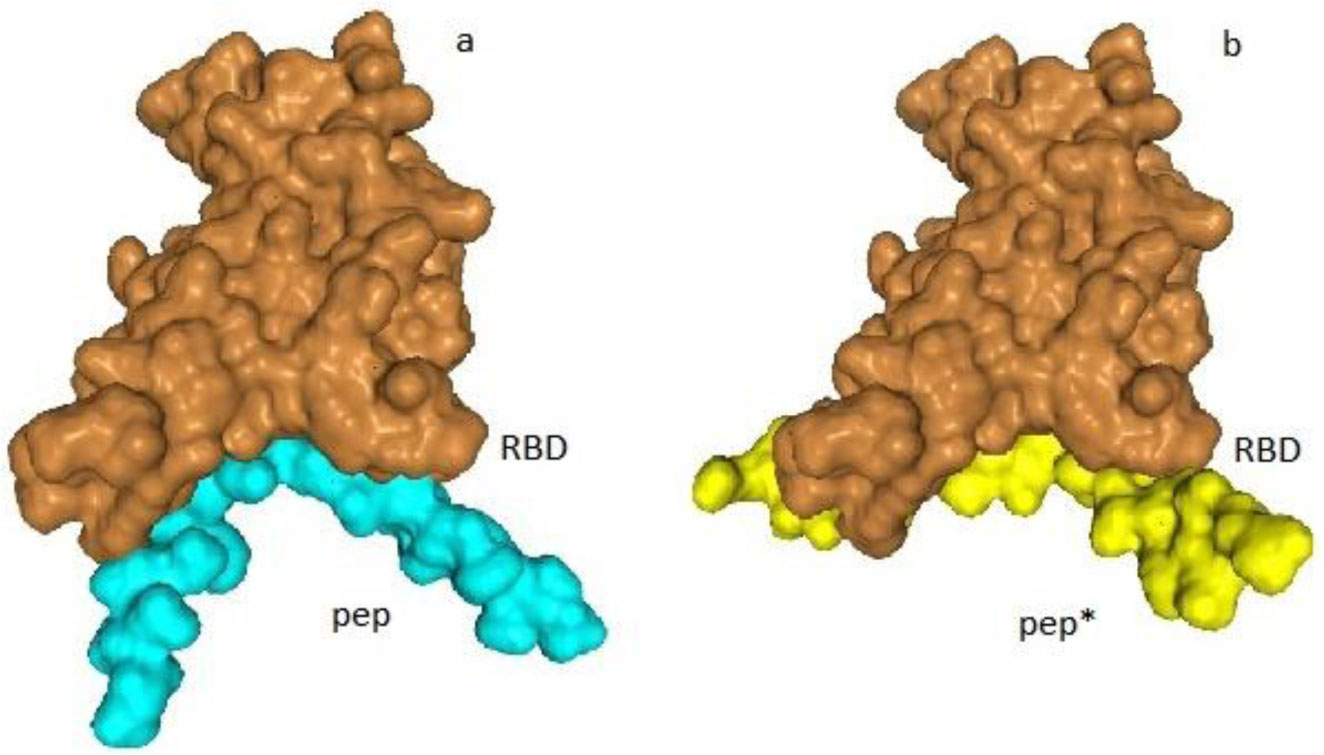
The charged amino acids in the spike protein of SARS-CoV-2 exhibit some specific distribution. In the RBD region of the S1 unit the positive charge dominates in the protein interior and the negative charge prevails on the surface exposed towards ACE2 receptor. The charged amino acids in the S2 region preceding heptad repeats of the spike protein of SARS-CoV-2, exhibit a central concentration, and the imbalance of a positive charge. The idea that both distributions of charged amino acids may, in an electrostatic manner, facilitate the coronavirus infection of the cell is presented. An evaluation of the virus-receptor binding energy, by docking the peptide resembling the human receptor site to the virus RBD, and the estimation of electromechanical deformation of the cell membrane by S2 during the prefusion process were included. Possible prevention of viral infection is suggested.
Citation: Piotr H. Pawłowski. Charged amino acids may promote coronavirus SARS-CoV-2 fusion with the host cell[J]. AIMS Biophysics, 2021, 8(1): 111-120. doi: 10.3934/biophy.2021008
The charged amino acids in the spike protein of SARS-CoV-2 exhibit some specific distribution. In the RBD region of the S1 unit the positive charge dominates in the protein interior and the negative charge prevails on the surface exposed towards ACE2 receptor. The charged amino acids in the S2 region preceding heptad repeats of the spike protein of SARS-CoV-2, exhibit a central concentration, and the imbalance of a positive charge. The idea that both distributions of charged amino acids may, in an electrostatic manner, facilitate the coronavirus infection of the cell is presented. An evaluation of the virus-receptor binding energy, by docking the peptide resembling the human receptor site to the virus RBD, and the estimation of electromechanical deformation of the cell membrane by S2 during the prefusion process were included. Possible prevention of viral infection is suggested.

| [1] |
Xia S, Liu M, Wang C, et al. (2020) Inhibition of SARS-CoV-2 (previously 2019-nCoV) infection by a highly potent pan-coronavirus fusion inhibitor targeting its spike protein that harbors a high capacity to mediate membrane fusion. Cell Res 30: 343-355. doi: 10.1038/s41422-020-0305-x

|
| [2] |
Millet JK, Whittaker GR (2018) Physiological and molecular triggers for SARS-CoV membrane fusion and entry into host cells. Virology 517: 3-8. doi: 10.1016/j.virol.2017.12.015
|
| [3] |
Lan J, Ge J, Yu J, et al. (2020) Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 581: 215-220. doi: 10.1038/s41586-020-2180-5
|
| [4] |
Lai AL, Millet JK, Daniel S, et al. (2017) The SARS-CoV fusion peptide forms an extended bipartite fusion platform that perturbs membrane order in a calcium-dependent manner. J Mol Biol 429: 3875-3892. doi: 10.1016/j.jmb.2017.10.017
|
| [5] |
Benarroch JM, Asally M (2020) The microbiologist's guide to membrane potential dynamics. Trends Microbiol 28: 304-314. doi: 10.1016/j.tim.2019.12.008
|
| [6] |
Yang J, Petitjean S, Derclaye S, et al. (2020) Molecular interaction and inhibition of SARS-CoV-2 binding to the ACE2 receptor. Nat Commun 4541. doi: 10.1038/s41467-020-18319-6
|
| [7] |
Zhou P, Jin B, Li H, et al. (2018) HPEPDOCK: a web server for blind peptide–protein docking based on a hierarchical algorithm. Nucleic Acids Res 46: W443-W450. doi: 10.1093/nar/gky357
|
| [8] |
Wong FC, Ong JH, Chai TT (2020) Identification of putative cell-entry-inhibitory peptides against SARS-CoV-2 from edible insects: an in silico study. EFood 1: 357-368. doi: 10.2991/efood.k.200918.002
|
| [9] |
Evans EA, Waugh R, Melnik L (1976) Elastic area compressibility modulus of red cell membrane. Biophys J 16: 585. doi: 10.1016/S0006-3495(76)85713-X
|
| [10] |
Cherstvy AG, Petrov EP (2014) Modeling DNA condensation on freestanding cationic lipid membranes. Phys Chem Chem Phys 16: 2020-2037. doi: 10.1039/C3CP53433B
|
| [11] |
Kumari S, Ye F, Podgornik R (2020) Ordering of adsorbed rigid rods mediated by the Boussinesq interaction on a soft substrate. J Chem Phys 153: 144905. doi: 10.1063/5.0022556
|
| [12] |
Lonappan A (2012) Novel method of detecting H1N1 using microwaves. J Biomed Sci Eng 5: 476-479. doi: 10.4236/jbise.2012.58060
|
| [13] |
Cherstvy AG, Thapa S, Wagner CE, et al. (2019) Non-Gaussian, non-ergodic, and non-Fickian diffusion of tracers in mucin hydrogels. Soft Matter 15: 2526-2551. doi: 10.1039/C8SM02096E
|
| [14] | Cantuti-Castelvetri L, Ojha R, Pedro LD, et al. (2020) Neuropilin-1 facilitates SARS-CoV-2 cell entry and provides a possible pathway into the central nervous system. BioRxiv preprint https://doi.org/10.1101/2020.06.07.137802. |
| [15] |
Xia S, Lan Q, Su S, et al. (2020) The role of furin cleavage site in SARS-CoV-2 spike protein-mediated membrane fusion in the presence or absence of trypsin. Signal Transduct TarTher 5: 1-3. doi: 10.1038/s41392-019-0089-y
|
| [16] |
Findlay EG, Currie SM, Davidson DJ (2013) Cationic host defence peptides: potential as antiviral therapeutics. Biodrugs 27: 479-493. doi: 10.1007/s40259-013-0039-0
|
| [17] |
Boas LCPV, Campos ML, Berlanda RLA, et al. (2019) Antiviral peptides as promising therapeutic drugs. Cell Mol Life Sci 76: 3525-3542. doi: 10.1007/s00018-019-03138-w
|
| [18] |
Straus MR, Tang T, Lai AL, et al. (2020) Ca2+ ions promote fusion of Middle East Respiratory Syndrome coronavirus with host cells and increase infectivity. J Virol 94: e00426-20. doi: 10.1128/JVI.00426-20
|
| [19] | Franke K (1954) Anionic water spray in the prophylaxis and treatment of bronchitis. Arch Phys Ther 6: 130-132. |
| [20] | Susuki T, Kobayashi N (2004) Plasmacluster ions inactivate an airborne corona virus: A world first verification research conducted jointly with the Kitasato Institute. Sharp Company, Press release 2004. |
| [21] |
Goswami DY (2003) Decontamination of ventilation systems using photocatalytic air cleaning technology. J Sol Energy Eng 125: 359-365. doi: 10.1115/1.1592540
|
 biophy-08-01-008-s001.pdf biophy-08-01-008-s001.pdf |
 |

Figures(8) / Tables(1)

Piotr H. Pawłowski. Charged amino acids may promote coronavirus SARS-CoV-2 fusion with the host cell[J]. AIMS Biophysics, 2021, 8(1): 111-120. doi: 10.3934/biophy.2021008







 DownLoad:
DownLoad: