Citation: Lars H. Wegner. Cotransport of water and solutes in plant membranes: The molecular basis, and physiological functions[J]. AIMS Biophysics, 2017, 4(2): 192-209. doi: 10.3934/biophy.2017.2.192

| [1] |
Poole RJ (1978) Energy coupling for membrane transport. Annu Rev Plant Physiol 29: 437–460. doi: 10.1146/annurev.pp.29.060178.002253

|
| [2] |
Williams LE, Lemoine R, Sauer N (2000) Sugar transporters in higher plants-a diversity of roles and complex regulation. Trends Plant Sci 5: 283–290. doi: 10.1016/S1360-1385(00)01681-2
|
| [3] |
Bush DR (1993) Proton-coupled sugar and amino acid transporters in plants. Annu Rev Plant Biol 44: 513–542. doi: 10.1146/annurev.pp.44.060193.002501
|
| [4] |
Wright EM, Turk E (2004) The sodium/glucose cotransport family SLC5. Pflüg Arch 447: 510–518. doi: 10.1007/s00424-003-1063-6
|
| [5] |
Raven JA, Doblin MA (2014) Active water transport in unicellular algae: where, why, and how. J Exp Bot 65: 6279–6292. doi: 10.1093/jxb/eru360
|
| [6] |
Wegner LH (2014) Root pressure and beyond: energetically uphill water transport into xylem vessels? J Exp Bot 65: 381–393. doi: 10.1093/jxb/ert391
|
| [7] |
Wegner LH (2015) A thermodynamic analysis of the feasibility of water secretion into xylem vessels against a water potential gradient. Funct Plant Biol 42: 828–835. doi: 10.1071/FP15077
|
| [8] | Wegner LH, (2015) Interplay of water and nutrient transport: A whole-plant perspective, In: Lüttge U, Beyschlag W, Progress in Botany, Springer, 109–141. |
| [9] |
Wegner LH (2017) A pump/leak model of growth: the biophysics of cell elongation in higher plants revisited. Funct Plant Biol 44: 185–197. doi: 10.1071/FP16184
|
| [10] |
Dainty J (1963) Water relations of plant cells. Adv Bot Res 1: 279–326. doi: 10.1016/S0065-2296(08)60183-4
|
| [11] | Zimmermann U, Steudle E (1978) Physical aspects of water relations of plant cells. Adv Bot Res 6: 45–117. |
| [12] |
Dainty J, Croghan PC, Fensom DS (1963) Electro-osmosis, with some applications to plant physiology. Can J Bot 41: 953–966. doi: 10.1139/b63-079
|
| [13] |
Tyerman SD, Schachtman DP (1992) The role of ion channels in plant nutrition and prospects for their genetic manipulation. Plant Soil 146: 137–144. doi: 10.1007/BF00012006
|
| [14] |
Rosenberg PA, Finkelstein A (1978) Interaction of ions and water in gramicidin A channels: streaming potentials across lipid bilayer membranes. J Gen Physiol 72: 327–340. doi: 10.1085/jgp.72.3.327
|
| [15] | Alcayaga C, Cecchi X, Alvarez O, et al. (1989) Streaming potential measurements in Ca2+-activated K+ channels from skeletal and smooth muscle. Coupling of ion and water fluxes. Biophys J 55: 367–371. |
| [16] |
Homblé F, Véry AA (1992) Coupling of water and potassium ions in K channels of the tonoplast of Chara. Biophys J 63: 996–999. doi: 10.1016/S0006-3495(92)81666-1
|
| [17] |
Levitt DG (1990) Streaming potential: Continuum expression applicable to very small nonuniform ion channels. J Chem Phys 92: 6953–6957. doi: 10.1063/1.458282
|
| [18] |
Barry PH, Hope AB (1969) Electro-osmosis in Chara and Nitella cells. Biochim Biophys Acta BBA-Biomembr 193: 124–128. doi: 10.1016/0005-2736(69)90065-0
|
| [19] |
Maurel C, Boursiac Y, Luu DT, et al. (2015) Aquaporins in plants. Physiol Rev 95: 1321–1358. doi: 10.1152/physrev.00008.2015
|
| [20] |
Chaumont F, Tyerman SD (2014) Aquaporins: highly regulated channels controlling plant water relations. Plant Physiol 164: 1600–1618. doi: 10.1104/pp.113.233791
|
| [21] |
Uehlein N, Lovisolo C, Siefritz F, et al. (2003) The tobacco aquaporin NtAQP1 is a membrane CO2 pore with physiological functions. Nature 425: 734–737. doi: 10.1038/nature02027
|
| [22] | Byrt CS, Zhao M, Kourghi M, et al. (2016) Non-selective cation channel activity of aquaporin AtPIP2; 1 regulated by Ca2+ and pH. Plant Cell Environ. Available from: http://onlinelibrary.wiley.com/doi/10.1111/pce.12832/full. |
| [23] |
Roberts SK, Tester M (1997) A patch clamp study of Na+ transport in maize roots. J Exp Bot 48: 431–440. doi: 10.1093/jxb/48.Special_Issue.431
|
| [24] |
Demidchik V, Tester M (2002) Sodium fluxes through nonselective cation channels in the plasma membrane of protoplasts from Arabidopsis roots. Plant Physiol 128: 379–387. doi: 10.1104/pp.010524
|
| [25] |
Yool AJ, Campbell EM (2012) Structure, function and translational relevance of aquaporin dual water and ion channels. Mol Aspects Med 33: 553–561. doi: 10.1016/j.mam.2012.02.001
|
| [26] |
Zeuthen T (2010) Water-transporting proteins. J Membr Biol 234: 57–73. doi: 10.1007/s00232-009-9216-y
|
| [27] |
Meinild AK, Klaerke DA, Loo DDF, et al. (1998) The human Na+-glucose cotransporter is a molecular water pump. J Physiol 508: 15–21. doi: 10.1111/j.1469-7793.1998.015br.x
|
| [28] |
Zeuthen T, MacAulay N (2012) Cotransport of water by Na+-K+-2Cl– cotransporters expressed in Xenopus oocytes: NKCC1 versus NKCC2. J Physiol 590: 1139–1154. doi: 10.1113/jphysiol.2011.226316
|
| [29] |
Loo DD, Hirayama BA, Meinild AK et al. (1999) Passive water and ion transport by cotransporters. J Physiol 518: 195–202. doi: 10.1111/j.1469-7793.1999.0195r.x
|
| [30] |
Colmenero-Flores JM, Martínez G, Gamba G, et al. (2007) Identification and functional characterization of cation-chloride cotransporters in plants. Plant J Cell Mol Biol 50: 278–292. doi: 10.1111/j.1365-313X.2007.03048.x
|
| [31] |
Kong XQ, Gao XH, Sun W, et al. (2011) Cloning and functional characterization of a cation-chloride cotransporter gene OsCCC1. Plant Mol Biol 75: 567–578. doi: 10.1007/s11103-011-9744-6
|
| [32] |
Loo DD, Zeuthen T, Chandy G, Wright EM (1996) Cotransport of water by the Na+/glucose cotransporter. Proc Natl Acad Sci 93: 13367–13370. doi: 10.1073/pnas.93.23.13367
|
| [33] | Zeuthen T, Gorraitz E, Her K, et al. (2016) Structural and functional significance of water permeation through cotransporters. Proc Natl Acad Sci 113: E6887–E6894. |
| [34] | Sauer N (2007) Molecular physiology of higher plant sucrose transporters. Febs Lett 581(12): 2309–2317. |
| [35] |
Geiger D (2011) Plant sucrose transporters from a biophysical point of view. Mol Plant 4: 395–406. doi: 10.1093/mp/ssr029
|
| [36] |
Chen LQ, Qu XQ, Hou BH, et al. (2012) Sucrose efflux mediated by SWEET proteins as a key step for phloem transport. Science 335: 207–211. doi: 10.1126/science.1213351
|
| [37] |
Oertli JJ (1966) Active water transport in plants. Physiol Plant 19: 809–817. doi: 10.1111/j.1399-3054.1966.tb07067.x
|
| [38] | Pickard WF (2003) The riddle of root pressure. II. Root exudation at extreme osmolalities. Funct Plant Biol 30: 135–141. |
| [39] | Zholkevich VN, (1991) Root pressure, In: Waisel Y, Eshel A, Kafkafi U, Plant roots, the hidden half Marcel-Dekker, New York, 589–603. |
| [40] |
Fricke W (2015) The significance of water co-transport for sustaining transpirational water flow in plants: a quantitative approach. J Exp Bot 66: 731–739. doi: 10.1093/jxb/eru466
|
| [41] | Henderson SW, Wege S, Qiu J, et al. (2015) Grapevine and Arabidopsis cation-chloride cotransporters localize to the golgi and trans-golgi network and indirectly influence long-distance ion transport and plant salt tolerance. Plant Physiol 169: 2215–2229. |
| [42] |
Boursiac Y, Boudet J, Postaire O, et al. (2008) Stimulus-induced downregulation of root water transport involves reactive oxygen species-activated cell signalling and plasma membrane intrinsic protein internalization. Plant J 56: 207–218. doi: 10.1111/j.1365-313X.2008.03594.x
|
| [43] |
Hirsch JR, Loo DD, Wright EM (1996) Regulation of Na+/glucose cotransporter expression by protein kinases in Xenopus laevis oocytes. J Biol Chem 271: 14740–14746. doi: 10.1074/jbc.271.25.14740
|
| [44] |
Benschop JJ, Mohammed S, O'Flaherty M, et al. (2007) Quantitative phosphoproteomics of early elicitor signaling in Arabidopsis. Mol Cell Proteomics 6: 1198–1214. doi: 10.1074/mcp.M600429-MCP200
|
| [45] |
MacRobbie EAC (1999) Vesicle trafficking: a role in trans-tonoplast ion movements? J Exp Bot 50: 925–934. doi: 10.1093/jxb/50.Special_Issue.925
|
| [46] |
Brüx A, Liu TY, Krebs M, Stierhof YD, et al. (2008) Reduced V-ATPase activity in the trans-Golgi network causes oxylipin-dependent hypocotyl growth inhibition in Arabidopsis. Plant Cell 20: 1088–1100. doi: 10.1105/tpc.108.058362
|
| [47] | Fricke W (2016) Water transport and energy. Plant Cell Environ. Available from: http://onlinelibrary.wiley.com/doi/10.1111/pce.12848/full. |
| [48] | Briskin DP, Reynolds-Niesman I (1991) Determination of H+/ATP stoichiometry for the plasma membrane H+-ATPase from red beet (Beta vulgaris L.) storage tissue. Plant Physiol 95: 242–250. |
| [49] |
Kaufmann I, Schulze-Till T, Schneider HU, et al. (2009) Functional repair of embolized vessels in maize roots after temporal drought stress, as demonstrated by magnetic resonance imaging. New Phytol 184: 245–256. doi: 10.1111/j.1469-8137.2009.02919.x
|
| [50] | Bentrup FW (2016) Water ascent in trees and lianas: the cohesion-tension theory revisited in the wake of Otto Renner. Protoplasma 254: 627–633. |
| [51] | Ryu J, Hwang BG, Lee SJ (2016) In vivo dynamic analysis of water refilling in embolized xylem vessels of intact Zea mays leaves. Ann Bot 118: 1033–1042. |
| [52] |
Morillon R, Liénard D, Chrispeels MJ, et al. (2001) Rapid movements of plants organs require solute-water cotransporters or contractile proteins. Plant Physiol 127: 720–723. doi: 10.1104/pp.010732
|
| [53] |
Tomos D, Pritchard J (1994) Biophysical and biochemical control of cell expansion in roots and leaves. J Exp Bot 45: 1721–1731. doi: 10.1093/jxb/45.Special_Issue.1721
|
| [54] |
Moshelion M, Becker D, Biela A, et al. (2002) Plasma Membrane aquaporins in the motor cells of samanea saman diurnal and circadian regulation. Plant Cell 14: 727–739. doi: 10.1105/tpc.010351
|
| [55] |
Lockhart JA (1965) An analysis of irreversible plant cell elongation. J Theor Biol 8: 264–275. doi: 10.1016/0022-5193(65)90077-9
|
| [56] |
Pritchard J (1994) The control of cell expansion in roots. New Phytol 127: 3–26. doi: 10.1111/j.1469-8137.1994.tb04255.x
|
| [57] |
Hukin D, Doering-Saad C, Thomas C, et al. (2002) Sensitivity of cell hydraulic conductivity to mercury is coincident with symplasmic isolation and expression of plasmalemma aquaporin genes in growing maize roots. Planta 215: 1047–1056. doi: 10.1007/s00425-002-0841-2
|
| [58] |
Ding L, Gao C, Li Y, et al. (2015) The enhanced drought tolerance of rice plants under ammonium is related to aquaporin (AQP). Plant Sci 234: 14–21. doi: 10.1016/j.plantsci.2015.01.016
|
| [59] | Zeuthen T (1996) Molecular mechanisms of water transport (RG Landes Compagny) Austin, Tx, USA. |
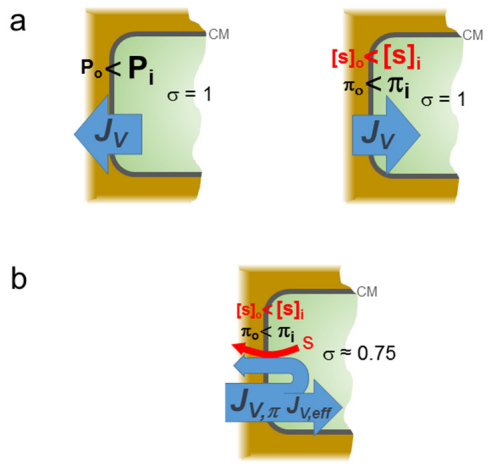
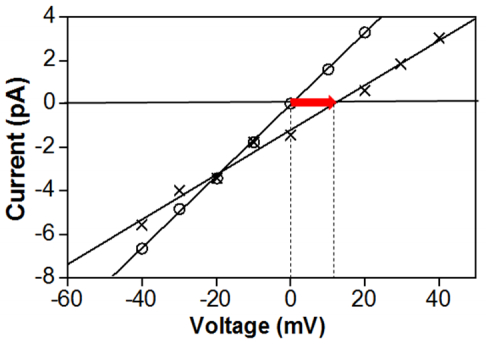
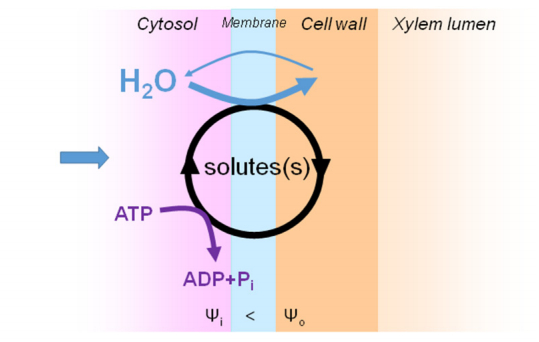
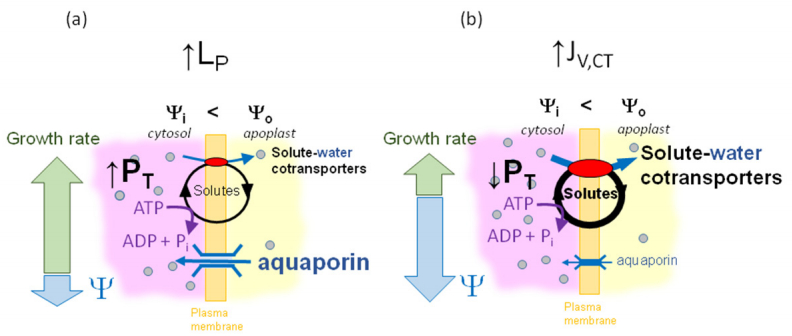
Figures(4)

Lars H. Wegner. Cotransport of water and solutes in plant membranes: The molecular basis, and physiological functions[J]. AIMS Biophysics, 2017, 4(2): 192-209. doi: 10.3934/biophy.2017.2.192







 DownLoad:
DownLoad: