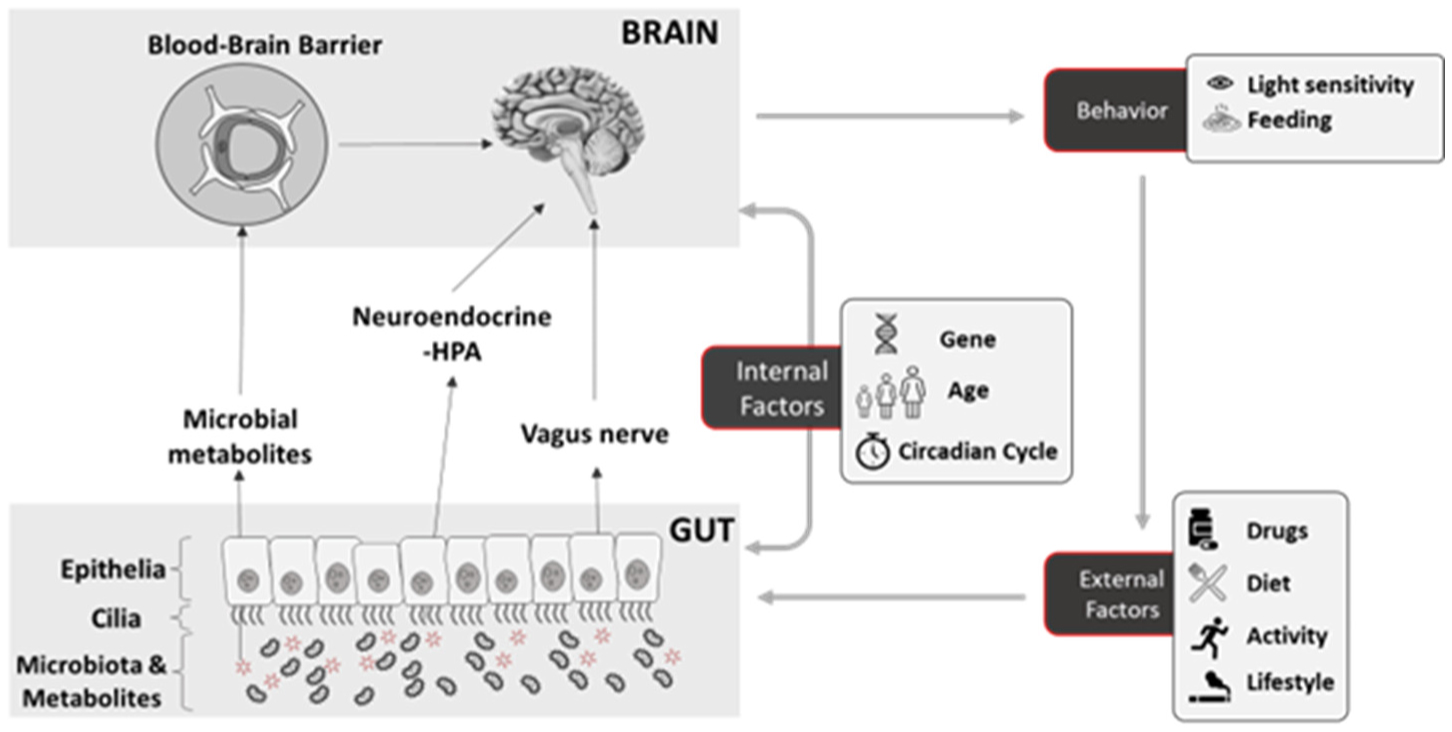
The brain-gut axis refers to the communication between the central nervous system and the gastrointestinal tract, with the gut microbiome playing a crucial role. While our understanding of the interaction between the gut microbiome and the host's physiology is still in its nascent stage, evidence suggests that the gut microbiota can indeed modulate host behavior. Understanding the specific mechanisms by which the gut microbiota community modulates the host's behavior remains the focus of present and future neuro-gastroenterology studies. This paper reviews several pieces of evidence from the literature on the impact of gut microbiota on host behavior across animal taxa. We explore the different pathways through which this modulation occurs, with the aim of deepening our understanding of the fascinating relationship between the gut microbiome and the central nervous system.
Citation: Temitope Awe, Ayoola Fasawe, Caleb Sawe, Adedayo Ogunware, Abdullahi Temitope Jamiu, Michael Allen. The modulatory role of gut microbiota on host behavior: exploring the interaction between the brain-gut axis and the neuroendocrine system[J]. AIMS Neuroscience, 2024, 11(1): 49-62. doi: 10.3934/Neuroscience.2024004
The brain-gut axis refers to the communication between the central nervous system and the gastrointestinal tract, with the gut microbiome playing a crucial role. While our understanding of the interaction between the gut microbiome and the host's physiology is still in its nascent stage, evidence suggests that the gut microbiota can indeed modulate host behavior. Understanding the specific mechanisms by which the gut microbiota community modulates the host's behavior remains the focus of present and future neuro-gastroenterology studies. This paper reviews several pieces of evidence from the literature on the impact of gut microbiota on host behavior across animal taxa. We explore the different pathways through which this modulation occurs, with the aim of deepening our understanding of the fascinating relationship between the gut microbiome and the central nervous system.

| [1] |
Das De T, Sharma P, Tevatiya S, et al. (2022) Bidirectional Microbiome-Gut-Brain-Axis Communication Influences Metabolic Switch-Associated Responses in the Mosquito Anopheles culicifacies. Cells 11: 24. https://doi.org/10.3390/cells11111798 
|
| [2] |
Sampson TR, Mazmanian SK (2015) Control of Brain Development, Function, and Behavior by the Microbiome. Cell Host Microbe 17: 565-576. https://doi.org/10.1016/j.chom.2015.04.011
|
| [3] |
Di Biase AR, Marasco G, Ravaioli F, et al. (2021) Gut microbiota signatures and clinical manifestations in celiac disease children at onset: a pilot study. J Gastroen Hepatol 36: 446-454. https://doi.org/10.1111/jgh.15183
|
| [4] |
Lynch JB, Gonzalez EL, Choy K, et al. (2023) Gut microbiota Turicibacter strains differentially modify bile acids and host lipids. Nat Commun 14: 15. https://doi.org/10.1038/s41467-023-39403-7
|
| [5] | Zhang Y, Baldyga K, Dong Y, et al. (2023) The association between gut microbiota and postoperative delirium in patients. Transl Psychiat 13. https://doi.org/10.1038/s41398-023-02450-1 |
| [6] |
Breton J, Tennoune N, Lucas N, et al. (2016) Gut Commensal E. coli Proteins Activate Host Satiety Pathways following Nutrient-Induced Bacterial Growth. Cell Metab 23: 324-334. https://doi.org/10.1016/j.cmet.2015.10.017
|
| [7] |
Leitao-Gonçalves R, Carvalho-Santos Z, Francisco AP, et al. (2017) Commensal bacteria and essential amino acids control food choice behavior and reproduction. Plos Biol 15: 29. https://doi.org/10.1371/journal.pbio.2000862
|
| [8] | Wiley NC, Dinan TG, Ross RP, et al. (2017) The microbiota-gut-brain axis as a key regulator of neural function and the stress response: Implications for human and animal health. J Anim Sci 95: 3225-3246. https://doi.org/10.2527/jas2016.1256 |
| [9] |
Johnson KVA, Foster KR (2018) Why does the microbiome affect behaviour?. Nat Rev Microbiol 16: 647-655. https://doi.org/10.1038/s41579-018-0014-3
|
| [10] |
Arnoriaga-Rodríguez M, Mayneris-Perxachs J, Burokas A, et al. (2020) Gut bacterial ClpB-like gene function is associated with decreased body weight and a characteristic microbiota profile. Microbiome 8: 10. https://doi.org/10.1186/s40168-020-00837-6
|
| [11] |
Henriques SF, Dhakan DB, Serra L, et al. (2020) Metabolic cross-feeding in imbalanced diets allows gut microbes to improve reproduction and alter host behaviour. Nat Commun 11: 15. https://doi.org/10.1038/s41467-020-18049-9
|
| [12] |
Han H, Yi B, Zhong RQ, et al. (2021) From gut microbiota to host appetite: gut microbiota-derived metabolites as key regulators. Microbiome 9: 16. https://doi.org/10.1186/s40168-021-01093-y
|
| [13] |
Dirksen P, Marsh SA, Braker I, et al. (2016) The native microbiome of the nematode Caenorhabditis elegans: gateway to a new host-microbiome model. Bmc Biol 14. https://doi.org/10.1186/s12915-016-0258-1
|
| [14] | Curtis JT, Assefa S, Francis A, et al. (2018) Fecal microbiota in the female prairie vole (Microtus ochrogaster). Plos One 13: 22. https://doi.org/10.1371/journal.pone.0190648 |
| [15] |
Turnbaugh PJ, Ley RE, Hamady M, et al. (2007) The Human Microbiome Project. Nature 449: 804-810. https://doi.org/10.1038/nature06244
|
| [16] |
Qin JJ, Li RQ, Raes J, et al. (2010) A human gut microbial gene catalogue established by metagenomic sequencing. Nature 464: 59-U70. https://doi.org/10.1038/nature08821
|
| [17] |
McKenney EA, Rodrigo A, Yoder AD (2015) Patterns of Gut Bacterial Colonization in Three Primate Species. Plos One 10. https://doi.org/10.1371/journal.pone.0124618
|
| [18] |
John GK, Mullin GE (2016) The Gut Microbiome and Obesity. Curr Oncol Rep 18: 7. https://doi.org/10.1007/s11912-016-0528-7
|
| [19] |
Cussotto S, Sandhu KV, Dinan TG, et al. (2018) The Neuroendocrinology of the Microbiota-Gut-Brain Axis: A Behavioural Perspective. Front Neuroendocrin 51: 80-101. https://doi.org/10.1016/j.yfrne.2018.04.002
|
| [20] | Richard ML, Sokol H (2019) The gut mycobiota: insights into analysis, environmental interactions and role in gastrointestinal diseases. Nat Rev Gastro Hepat 16: 331-345. https://doi.org/10.1038/s41575-019-0121-2 |
| [21] | Carabotti M, Scirocco A, Maselli MA, et al. (2015) The gut-brain axis: interactions between enteric microbiota, central and enteric nervous systems. Ann Gastroenterol 28: 7. |
| [22] |
Ortega MA, Alvarez-Mon MA, García-Montero C, et al. (2023) Microbiota-gut-brain axis mechanisms in the complex network of bipolar disorders: potential clinical implications and translational opportunities. Mol Psychiatr 28: 2645-2673. https://doi.org/10.1038/s41380-023-01964-w
|
| [23] |
Kim DH, Flavell SW (2020) Host-microbe interactions and the behavior of Caenorhabditis elegans. J Neurogenet 34: 500-509. https://doi.org/10.1080/01677063.2020.1802724
|
| [24] | Trevelline B, Kohl K (2023) The gut microbiome influences host diet selection behavior. Integr Comp Biol 63: S463-S463. |
| [25] |
Ousey J, Boktor JC, Mazmanian SK (2023) Gut microbiota suppress feeding induced by palatable foods. Curr Biol 33: 147-157.E7. https://doi.org/10.1016/j.cub.2022.10.066
|
| [26] |
Breton J, Tirelle P, Hasanat S, et al. (2021) Gut microbiota alteration in a mouse model of Anorexia Nervosa. Clin Nutr 40: 181-189. https://doi.org/10.1016/j.clnu.2020.05.002
|
| [27] |
Mondot S, Lachkar L, Doré J, et al. (2022) Roseburia, a decreased bacterial taxon in the gut microbiota of patients suffering from anorexia nervosa. Eur J Clin Nutr 76: 1486-1489. https://doi.org/10.1038/s41430-022-01116-3
|
| [28] |
Fan Y, Stoving RK, Ibraim SB, et al. (2023) The gut microbiota contributes to the pathogenesis of anorexia nervosa in humans and mice. Nat Microbiol 8: 787-802. https://doi.org/10.1038/s41564-023-01355-5
|
| [29] |
Labbadia J, Morimoto RI (2015) Repression of the Heat Shock Response Is a Programmed Event at the Onset of Reproduction. Mol Cell 59: 639-650. https://doi.org/10.1016/j.molcel.2015.06.027
|
| [30] |
Plagens RN, Mossiah I, Guisbert KSK, et al. (2021) Chronic temperature stress inhibits reproduction and disrupts endocytosis via chaperone titration in Caenorhabditis elegans. Bmc Biol 19: 14. https://doi.org/10.1186/s12915-021-01008-1
|
| [31] |
Mindus C, van Staaveren N, Bharwani A, et al. (2021) Ingestion of Lactobacillus rhamnosus modulates chronic stress-induced feather pecking in chickens. Sci Rep 11: 14. https://doi.org/10.1038/s41598-021-96615-x
|
| [32] |
Hsiao EY, McBride SW, Hsien S, et al. (2013) Microbiota Modulate Behavioral and Physiological Abnormalities Associated with Neurodevelopmental Disorders. Cell 155: 1451-1463. https://doi.org/10.1016/j.cell.2013.11.024
|
| [33] |
Buffington SA, Di Prisco GV, Auchtung TA, et al. (2016) Microbial Reconstitution Reverses Maternal Diet-Induced Social and Synaptic Deficits in Offspring. Cell 165: 1762-1775. https://doi.org/10.1016/j.cell.2016.06.001
|
| [34] |
Zhu F, Guo RJ, Wang W, et al. (2020) Transplantation of microbiota from drug-free patients with schizophrenia causes schizophrenia-like abnormal behaviors and dysregulated kynurenine metabolism in mice. Mol Psychiatr 25: 2905-2918. https://doi.org/10.1038/s41380-019-0475-4
|
| [35] |
Gualtieri P, Marchetti M, Cioccoloni G, et al. (2020) Psychobiotics Regulate the Anxiety Symptoms in Carriers of Allele A of IL-1β Gene: A Randomized, Placebo-Controlled Clinical Trial. Mediat Inflamm 2020: 11. https://doi.org/10.1155/2020/2346126
|
| [36] |
Dao VH, Hoang LB, Trinh TO, et al. (2021) Psychobiotics for Patients with Chronic Gastrointestinal Disorders Having Anxiety or Depression Symptoms. J Multidiscip Health 14: 1395-1402. https://doi.org/10.2147/JMDH.S312316
|
| [37] | Gohari E JL, Shafazand S (2019) The Effects of Altered Microbiome on Caenorhabditis elegans Egg Laying Behavior. J Emerging Investigators 2: 5. https://doi.org/10.59720/19-016 |
| [38] |
O'Donnell MP, Fox BW, Chao PH, et al. (2020) A neurotransmitter produced by gut bacteria modulates host sensory behaviour. Nature 583: 415-420. https://doi.org/10.1038/s41586-020-2395-5
|
| [39] | Hernandez-Caballero I, Garcia-Longoria L, Gomez-Mestre I, et al. (2022) The Adaptive Host Manipulation Hypothesis: Parasites Modify the Behaviour, Morphology, and Physiology of Amphibians. Diversity-Basel 14: 18. https://doi.org/10.3390/d14090739 |
| [40] |
Cox LM, Weiner HL (2018) Microbiota Signaling Pathways that Influence Neurologic Disease. Neurotherapeutics 15: 135-145. https://doi.org/10.1007/s13311-017-0598-8
|
| [41] |
Caspani G, Swann J (2019) Small talk: microbial metabolites involved in the signaling from microbiota to brain. Curr Opin Pharmacol 48: 99-106. https://doi.org/10.1016/j.coph.2019.08.001
|
| [42] |
Bercik P, Park AJ, Sinclair D, et al. (2011) The anxiolytic effect of Bifidobacterium longum NCC3001 involves vagal pathways for gut-brain communication. Neurogastroent Motil 23: 1132-E1544. https://doi.org/10.1111/j.1365-2982.2011.01796.x
|
| [43] |
Bravo JA, Forsythe P, Chew MV, et al. (2011) Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. P Natl Acad Sci USA 108: 16050-16055. https://doi.org/10.1073/pnas.1102999108
|
| [44] |
Siopi E, Galerne M, Rivagorda M, et al. (2023) Gut microbiota changes require vagus nerve integrity to promote depressive-like behaviors in mice. Mol Psychiatr 28: 3002-3012. https://doi.org/10.1038/s41380-023-02071-6
|
| [45] |
Braniste V, Al-Asmakh M, Kowal C, et al. (2014) The gut microbiota influences blood-brain barrier permeability in mice. Sci Transl Med 6. https://doi.org/10.1126/scitranslmed.3009759
|
| [46] |
Tang W, Zhu HF, Feng YM, et al. (2020) The Impact of Gut Microbiota Disorders on the Blood-Brain Barrier. Infect Drug Resist 13: 3351-3363. https://doi.org/10.2147/IDR.S254403
|
| [47] |
Luczynski P, Whelan SO, O'Sullivan C, et al. (2016) Adult microbiota-deficient mice have distinct dendritic morphological changes: differential effects in the amygdala and hippocampus. Eur J Neurosci 44: 2654-2666. https://doi.org/10.1111/ejn.13291
|
| [48] |
Dinan TG, Cryan JF (2017) Gut instincts: microbiota as a key regulator of brain development, ageing and neurodegeneration. J Physiol-London 595: 489-503. https://doi.org/10.1113/JP273106
|
| [49] |
Mosaferi B, Jand Y, Salari AA (2021) Antibiotic-induced gut microbiota depletion from early adolescence exacerbates spatial but not recognition memory impairment in adult male C57BL/6 mice with Alzheimer-like disease. Brain Res Bull 176: 8-17. https://doi.org/10.1016/j.brainresbull.2021.08.004
|
| [50] | Tang W, Meng ZY, Li N, et al. (2021) Roles of Gut Microbiota in the Regulation of Hippocampal Plasticity, Inflammation, and Hippocampus-Dependent Behaviors. Front Cell Infect Mi 10: 15. https://doi.org/10.3389/fcimb.2020.611014 |
| [51] |
Silva YP, Bernardi A, Frozza RL (2020) The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front Endocrinol 11: 14. https://doi.org/10.3389/fendo.2020.00025
|
| [52] |
Torres-Fuentes C, Golubeva AV, Zhdanov AV, et al. (2019) Short-chain fatty acids and microbiota metabolites attenuate ghrelin receptor signaling. Faseb J 33: 13546-13559. https://doi.org/10.1096/fj.201901433R
|
| [53] |
Frost G, Sleeth ML, Sahuri-Arisoylu M, et al. (2014) The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat Commun 5. https://doi.org/10.1038/ncomms4611
|
| [54] |
Yano JM, Yu K, Donaldson GP, et al. (2015) Indigenous Bacteria from the Gut Microbiota Regulate Host Serotonin Biosynthesis (vol 161, pg 264, 2015). Cell 163: 258-258. https://doi.org/10.1016/j.cell.2015.09.017
|
| [55] |
Clarke G, Grenham S, Scully P, et al. (2013) The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol Psychiatr 18: 666-673. https://doi.org/10.1038/mp.2012.77
|
| [56] |
Heijtza RD, Wang SG, Anuar F, et al. (2011) Normal gut microbiota modulates brain development and behavior. P Natl Acad Sci USA 108: 3047-3052. https://doi.org/10.1073/pnas.1010529108
|
| [57] |
Hoban AE, Stilling RM, Ryan FJ, et al. (2016) Regulation of prefrontal cortex myelination by the microbiota. Transl Psychiat 6. https://doi.org/10.1038/tp.2016.42
|
| [58] |
Larroya-García A, Navas-Carrillo D, Orenes-Piñero E (2019) Impact of gut microbiota on neurological diseases: Diet composition and novel treatments. Crit Rev Food Sci 59: 3102-3116. https://doi.org/10.1080/10408398.2018.1484340
|
| [59] |
Halmos T, Suba I (2016) Physiological patterns of intestinal microbiota. The role of dysbacteriosis in obesity, insulin resistance, diabetes and metabolic syndrome. Orvosi Hetilap 157: 13-22. https://doi.org/10.1556/650.2015.30296
|
| [60] |
Zhang CC, Björkman A, Cai K, et al. (2018) Impact of a 3-Months Vegetarian Diet on the Gut Microbiota and Immune Repertoire. Front Immunol 9: 13. https://doi.org/10.3389/fimmu.2018.00908
|
| [61] |
Pickel L, Lee JH, Maughan H, et al. (2022) Circadian rhythms in metabolic organs and the microbiota during acute fasting in mice. Physiol Rep 10: 18. https://doi.org/10.14814/phy2.15393
|
| [62] |
Qin YW, Havulinna AS, Liu Y, et al. (2022) Combined effects of host genetics and diet on human gut microbiota and incident disease in a single population cohort. Nat Genet 54: 134-142. https://doi.org/10.1038/s41588-021-00991-z
|
| [63] |
Sekirov I, Russell SL, Antunes LCM, et al. (2010) Gut Microbiota in Health and Disease. Physiol Rev 90: 859-904. https://doi.org/10.1152/physrev.00045.2009
|
| [64] |
Ticinesi A, Milani C, Lauretani F, et al. (2017) Gut microbiota composition is associated with polypharmacy in elderly hospitalized patients. Sci Rep 7: 11. https://doi.org/10.1038/s41598-017-10734-y
|
| [65] |
Clarke SF, Murphy EF, O'Sullivan O, et al. (2014) Exercise and associated dietary extremes impact on gut microbial diversity. Gut 63: 1913-1920. https://doi.org/10.1136/gutjnl-2013-306541
|
| [66] |
de Sire A, de Sire R, Petito V, et al. (2020) Gut-Joint Axis: The Role of Physical Exercise on Gut Microbiota Modulation in Older People with Osteoarthritis. Nutrients 12: 14. https://doi.org/10.3390/nu12020574
|
| [67] |
Gurwara S, Dai AN, Ajami NJ, et al. (2020) Alcohol use alters the colonicmucosa-associated gut microbiota in humans. Nutr Res 83: 119-128. https://doi.org/10.1016/j.nutres.2020.09.004
|
| [68] |
Thaiss CA, Levy M, Korem T, et al. (2016) Microbiota Diumal Rhythmicity Programs Host Transcriptome Oscillations. Cell 167: 1495-1510.E12. https://doi.org/10.1016/j.cell.2016.11.003
|
| [69] |
Nawroth JC, Guo HL, Koch E, et al. (2017) Motile cilia create fluid-mechanical microhabitats for the active recruitment of the host microbiome. P Natl Acad Sci USA 114: 9510-9516. https://doi.org/10.1073/pnas.1706926114
|
| [70] |
Hashimoto K (2023) Emerging role of the host microbiome in neuropsychiatric disorders: overview and future directions. Mol Psychiatr 28: 13. https://doi.org/10.1038/s41380-023-02287-6
|
| [71] |
Kennedy EA, King KY, Baldridge MT (2018) Mouse microbiota models: comparing germ-free mice and antibiotics treatment as tools for modifying gut bacteria. Front Physiol 9. https://doi.org/10.3389/fphys.2018.01534
|
| [72] | Wan JUN, Kang YJ, Chen FEI, et al. (2013) Application of an antibiotic crescent-shaped polymethylmethacrylate strut in thoracic vertebral tuberculosis. Turk Neurosurg 23: 6. |
| [73] |
Ichinohe T, Pang IK, Kumamoto Y, et al. (2011) Microbiota regulates immune defense against respiratory tract influenza A virus infection. P Natl Acad Sci USA 108: 5354-5359. https://doi.org/10.1073/pnas.1019378108
|
Figures(1) / Tables(1)

Temitope Awe, Ayoola Fasawe, Caleb Sawe, Adedayo Ogunware, Abdullahi Temitope Jamiu, Michael Allen. The modulatory role of gut microbiota on host behavior: exploring the interaction between the brain-gut axis and the neuroendocrine system[J]. AIMS Neuroscience, 2024, 11(1): 49-62. doi: 10.3934/Neuroscience.2024004







 DownLoad:
DownLoad: