Citation: Masatoshi Takita, Yumi Izawa-Sugaya. Neurocircuit differences between memory traces of persistent hypoactivity and freezing following fear conditioning among the amygdala, hippocampus, and prefrontal cortex[J]. AIMS Neuroscience, 2021, 8(2): 195-211. doi: 10.3934/Neuroscience.2021010

| [1] |
LeDoux JE (2000) Emotion circuits in the brain. Annu Rev Neurosci 23: 155-184. doi: 10.1146/annurev.neuro.23.1.155

|
| [2] |
Maren S (2001) Neurobiology of Pavlovian fear conditioning. Annu Rev Neurosci 24: 897-931. doi: 10.1146/annurev.neuro.24.1.897
|
| [3] |
Quirk GJ, Mueller D (2008) Neural mechanisms of extinction learning and retrieval. Neuropsychopharmacology 33: 56-72. doi: 10.1038/sj.npp.1301555
|
| [4] |
McSweeney FK, Swindell S (2002) Common processes may contribute to extinction and habituation. J Gen Psychol 129: 364-400. doi: 10.1080/00221300209602103
|
| [5] |
Thompson RF, Spencer WA (1966) Habituation: a model phenomenon for the study of neuronal substrates of behavior. Psychol Rev 73: 16-43. doi: 10.1037/h0022681
|
| [6] |
Phillips RG, LeDoux JE (1992) Differential contribution of amygdala and hippocampus to cued and contextual fear conditioning. Behav Neurosci 106: 274-285. doi: 10.1037/0735-7044.106.2.274
|
| [7] |
Kestenbaum R, Celman SA (1995) Preschool children's identification and understanding of mixed emotions. Cognit Dev 10: 443-458. doi: 10.1016/0885-2014(95)90006-3
|
| [8] |
Scrimin S, Moscardino U, Capello F, et al. (2009) Recognition of facial expressions of mixed emotions in school-age children exposed to terrorism. Dev Psychol 45: 1341-1352. doi: 10.1037/a0016689
|
| [9] |
Shin LM, Rauch SL, Pitman RK (2006) Amygdala, medial prefrontal cortex, and hippocampal function in PTSD. Ann N Y Acad Sci 1071: 67-79. doi: 10.1196/annals.1364.007
|
| [10] |
Bandelow B, Zohar J, Hollander E, et al. (2002) World Federation of Societies of Biological Psychiatry (WFSBP) guidelines for the pharmacological treatment of anxiety, obsessive-compulsive and post-traumatic stress disorders-first revision. World J Biol Psychiatry 9: 248-312. doi: 10.1080/15622970802465807
|
| [11] |
Corley MJ, Caruso MJ, Takahashi LK (2012) Stress-induced enhancement of fear conditioning and sensitization facilitates extinction-resistant and habituation-resistant fear behaviors in a novel animal model of posttraumatic stress disorder. Physiol Behav 105: 408-416. doi: 10.1016/j.physbeh.2011.08.037
|
| [12] |
Wilensky AE, Schafe GE, Kristensen MP, et al. (2006) Rethinking the fear circuit: the central nucleus of the amygdala is required for the acquisition, consolidation, and expression of Pavlovian fear conditioning. J Neurosci 26: 12387-12396. doi: 10.1523/JNEUROSCI.4316-06.2006
|
| [13] |
Bast T, Zhang WN, Feldon J (2001) The ventral hippocampus and fear conditioning in rats. Different anterograde amnesias of fear after tetrodotoxin inactivation and infusion of the GABA(A) agonist muscimol. Exp Brain Res 139: 39-52. doi: 10.1007/s002210100746
|
| [14] |
Maren S (1999) Long-term potentiation in the amygdala: a mechanism for emotional learning and memory. Trends Neurosci 22: 561-567. doi: 10.1016/S0166-2236(99)01465-4
|
| [15] |
Richmond MA, Yee BK, Pouzet B, et al. (1999) Dissociating context and space within the hippocampus: effects of complete, dorsal, and ventral excitotoxic hippocampal lesions on conditioned freezing and spatial learning. Behav Neurosci 113: 1189-1203. doi: 10.1037/0735-7044.113.6.1189
|
| [16] |
Lacroix L, Spinelli S, Heidbreder CA, et al. (2000) Differential role of the medial and lateral prefrontal cortices in fear and anxiety. Behav Neurosci 114: 1119-1130. doi: 10.1037/0735-7044.114.6.1119
|
| [17] |
Morgan MA, LeDoux JE (1995) Differential contribution of dorsal and ventral medial prefrontal cortex to the acquisition and extinction of conditioned fear in rats. Behav Neurosci 109: 681-688. doi: 10.1037/0735-7044.109.4.681
|
| [18] |
Morgan MA, Romanski LM, LeDoux JE (1993) Extinction of emotional learning: contribution of medial prefrontal cortex. Neurosci Lett 163: 109-113. doi: 10.1016/0304-3940(93)90241-C
|
| [19] |
Quirk GJ, Russo GK, Barron JL, et al. (2000) The role of ventromedial prefrontal cortex in the recovery of extinguished fear. J Neurosci 20: 6225-6231. doi: 10.1523/JNEUROSCI.20-16-06225.2000
|
| [20] |
Canteras NS, Swanson LW (1992) Projections of the ventral subiculum to the amygdala, septum, and hypothalamus: a PHAL anterograde tract-tracing study in the rat. J Comp Neurol 324: 180-194. doi: 10.1002/cne.903240204
|
| [21] |
Krettek JE, Price JL (1974) A direct input from the amygdala to the thalamus and the cerebral cortex. Brain Res 67: 169-174. doi: 10.1016/0006-8993(74)90309-6
|
| [22] |
Krettek JE, Price JL (1977) Projections from the amygdaloid complex to the cerebral cortex and thalamus in the rat and cat. J Comp Neurol 172: 687-722. doi: 10.1002/cne.901720408
|
| [23] |
Ottersen OP (1982) Connections of the amygdala of the rat. IV: Corticoamygdaloid and intraamygdaloid connections as studied with axonal transport of horseradish peroxidase. J Comp Neurol 205: 30-48. doi: 10.1002/cne.902050104
|
| [24] | Pitkänen A (2000) Connectivity of the rat amygdaloid complex. The Amygdala: A Functional Analysis New York: Oxford University Press, 31-115. |
| [25] |
Shi CJ, Cassell MD (1999) Perirhinal cortex projections to the amygdaloid complex and hippocampal formation in the rat. J Comp Neurol 406: 299-328. doi: 10.1002/(SICI)1096-9861(19990412)406:3<299::AID-CNE2>3.0.CO;2-9
|
| [26] |
Krettek JE, Price JL (1974) Projections from the amygdala to the perirhinal and entorhinal cortices and the subiculum. Brain Res 17: 150-154. doi: 10.1016/0006-8993(74)90199-1
|
| [27] |
Krettek JE, Price JL (1977) Projections from the amygdaloid complex and adjacent olfactory structures to the entorhinal cortex and to the subiculum in the rat and cat. J Comp Neurol 2: 723-752. doi: 10.1002/cne.901720409
|
| [28] |
Petrovich GD, Risold PY, Swanson LW (1996) Organization of projections from the basomedial nucleus of the amygdala: a PHAL study in the rat. J Comp Neurol 374: 387-420. doi: 10.1002/(SICI)1096-9861(19961021)374:3<387::AID-CNE6>3.0.CO;2-Y
|
| [29] |
Sarter M, Markowitsch HJ (1984) Collateral innervation of the medial and lateral prefrontal cortex by amygdaloid, thalamic, and brain-stem neurons. J Comp Neurol 224: 445-460. doi: 10.1002/cne.902240312
|
| [30] |
Vertes RP (2004) Differential projections of the infralimbic and prelimbic cortex in the rat. Synapse 51: 32-58. doi: 10.1002/syn.10279
|
| [31] |
McDonald AJ (1987) Organization of amygdaloid projections to the mediodorsal thalamus and prefrontal cortex: a fluorescence retrograde transport study in the rat. J Comp Neurol 262: 46-58. doi: 10.1002/cne.902620105
|
| [32] |
McDonald AJ, Mascagni F, Guo L (1996) Projections of the medial and lateral prefrontal cortices to the amygdala: a Phaseolus vulgaris leucoagglutinin study in the rat. Neuroscience 71: 55-75. doi: 10.1016/0306-4522(95)00417-3
|
| [33] |
Ferino F, Thierry AM, Glowinski J (1987) Anatomical and electrophysiological evidence for a direct projection from Ammon's horn to the medial prefrontal cortex in the rat. Exp Brain Res 65: 421-426. doi: 10.1007/BF00236315
|
| [34] |
Swanson LW (1981) A direct projection from Ammon's horn to prefrontal cortex in the rat. Brain Res 217: 150-154. doi: 10.1016/0006-8993(81)90192-X
|
| [35] |
Takita M, Fujiwara SE, Izaki Y (2013) Functional structure of the intermediate and ventral hippocampo-prefrontal pathway in the prefrontal convergent system. J Physiol Paris 107: 441-447. doi: 10.1016/j.jphysparis.2013.05.002
|
| [36] |
Herry C, Ferraguti F, Singewald N, et al. (2010) Neuronal circuits of fear extinction. Eur J Neurosci 31: 599-612. doi: 10.1111/j.1460-9568.2010.07101.x
|
| [37] |
Canteras NS, Resstel LB, Bertoglio LJ, et al. (2010) Neuroanatomy of anxiety. Curr Top Behav Neurosci 2: 77-96. doi: 10.1007/7854_2009_7
|
| [38] |
Singewald N, Holmes A (2019) Rodent models of impaired fear extinction. Psychopharmacology (Berl) 236: 21-32. doi: 10.1007/s00213-018-5054-x
|
| [39] |
Izaki Y, Takita M, Akema T (2008) Specific role of the posterior dorsal hippocampus-prefrontal cortex in short-term working memory. Eur J Neurosci 27: 3029-3034. doi: 10.1111/j.1460-9568.2008.06284.x
|
| [40] | Paxinos G, Watson G (1998) The rat brain in stereotaxic coordinates Sydney: Academic. |
| [41] |
Floresco SB, Seamans JK, Phillips AG (1997) Selective roles for hippocampal, prefrontal cortical, and ventral striatal circuits in radial-arm maze tasks with or without a delay. J Neurosci 17: 1880-1890. doi: 10.1523/JNEUROSCI.17-05-01880.1997
|
| [42] |
Bouton ME (2004) Context and behavioral processes in extinction. Learn Mem 11: 485-494. doi: 10.1101/lm.78804
|
| [43] |
Dudai Y (2006) Reconsolidation: the advantage of being refocused. Curr Opin Neurobiol 16: 174-178. doi: 10.1016/j.conb.2006.03.010
|
| [44] |
Dudai Y, Eisenberg M (2004) Rites of passage of the engram: reconsolidation and the lingering consolidation hypothesis. Neuron 44: 93-100. doi: 10.1016/j.neuron.2004.09.003
|
| [45] |
Eysenck HJ (1968) A theory of the incubation of anxiety-fear responses. Behav Res Ther 6: 309-321. doi: 10.1016/0005-7967(68)90064-8
|
| [46] | Houston FP, Stevenson GD, McNaughton BL, et al. (1999) Effects of age on the generalization and incubation of memory in the F344 rat. Learn Mem 6: 111-119. |
| [47] |
Rapaport MH, Clary C, Fayyad R, et al. (2005) Quality-of-life impairment in depressive and anxiety disorders. Am J Psychiatry 162: 1171-1178. doi: 10.1176/appi.ajp.162.6.1171
|
| [48] |
Bouton ME, Bolles RC (1980) Conditioned Fear Assessed by Freezing and by the Suppression of 3 Different Baselines. Anim Learn Behav 8: 429-434. doi: 10.3758/BF03199629
|
| [49] |
Amorapanth P, Nader K, LeDoux JE (1999) Lesions of periaqueductal gray dissociate-conditioned freezing from conditioned suppression behavior in rats. Learn Mem 6: 491-499. doi: 10.1101/lm.6.5.491
|
| [50] |
Kim JJ, Rison RA, Fanselow MS (1993) Effects of amygdala, hippocampus, and periaqueductal gray lesions on short- and long-term contextual fear. Behav Neurosci 107: 1093-1098. doi: 10.1037/0735-7044.107.6.1093
|
| [51] |
Vianna DM, Graeff FG, Landeira-Fernandez J, et al. (2001) Lesion of the ventral periaqueductal gray reduces conditioned fear but does not change freezing induced by stimulation of the dorsal periaqueductal gray. Learn Mem 8: 164-169. doi: 10.1101/lm.36101
|
| [52] |
Seidenbecher T, Laxmi TR, Stork O, et al. (2003) Amygdalar and hippocampal theta rhythm synchronization during fear memory retrieval. Science 301: 846-850. doi: 10.1126/science.1085818
|
| [53] |
Arnsten AFT (2009) Stress signalling pathways that impair prefrontal cortex structure and function. Nat Rev Neurosci 10: 410-422. doi: 10.1038/nrn2648
|
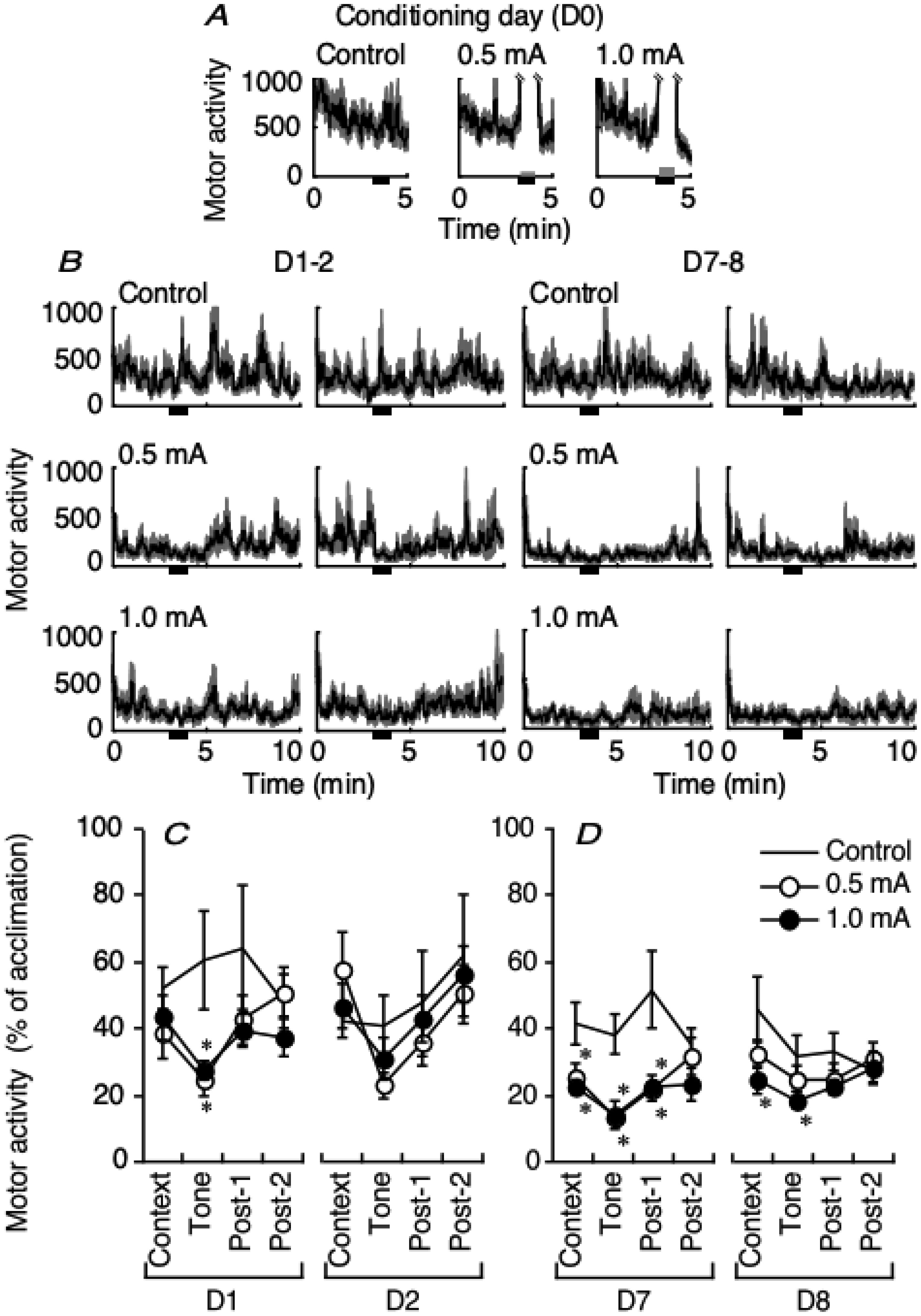
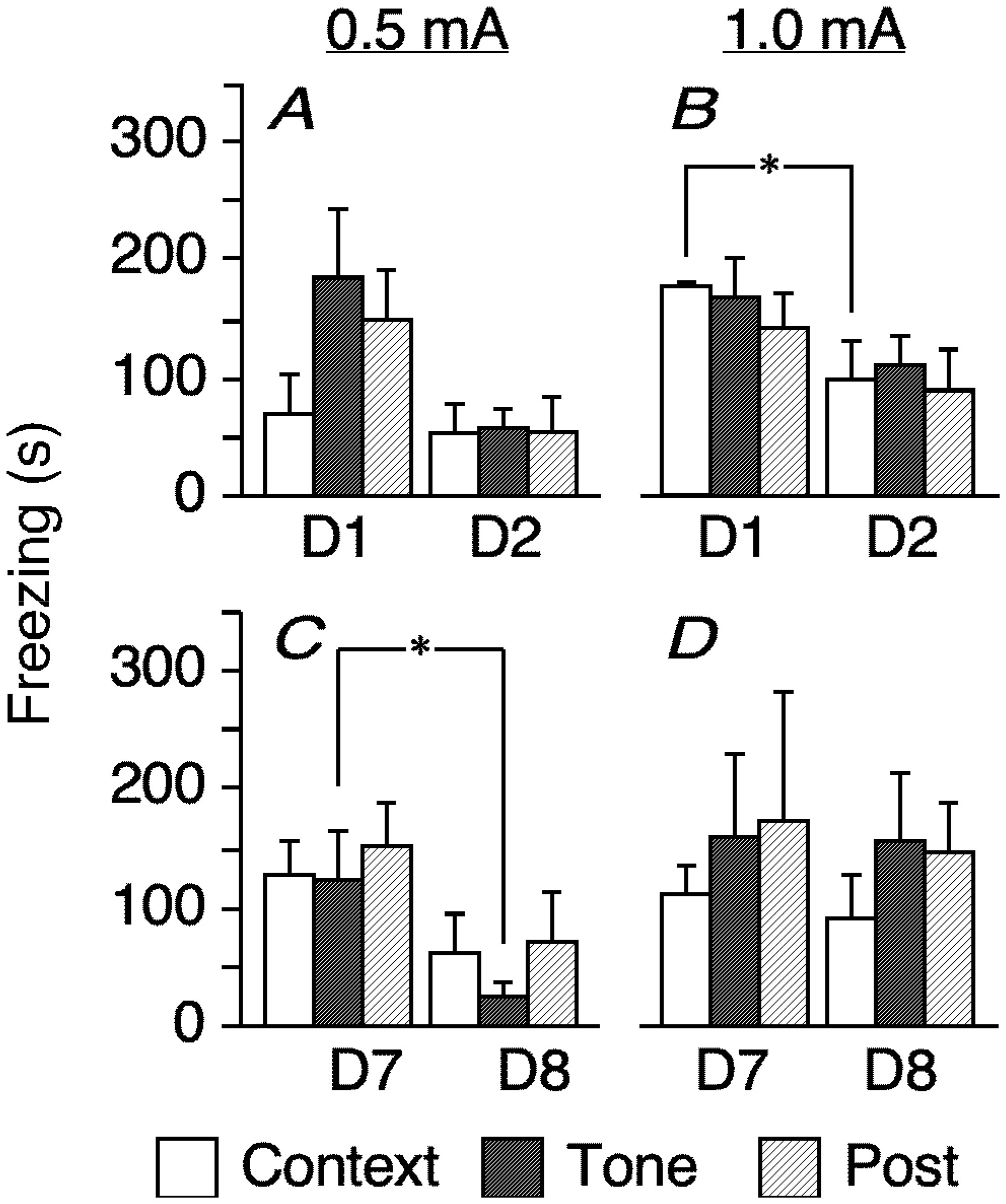
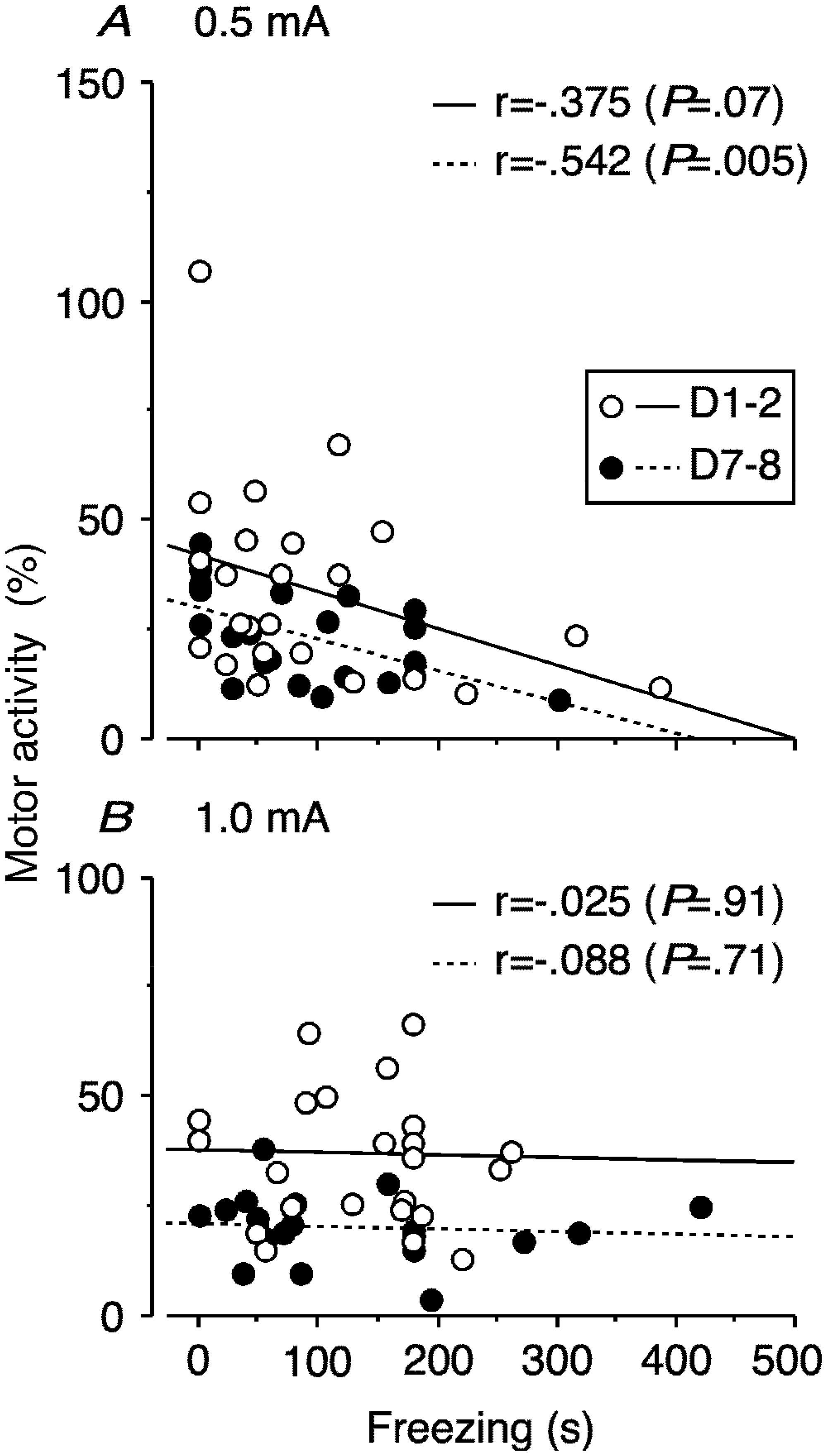
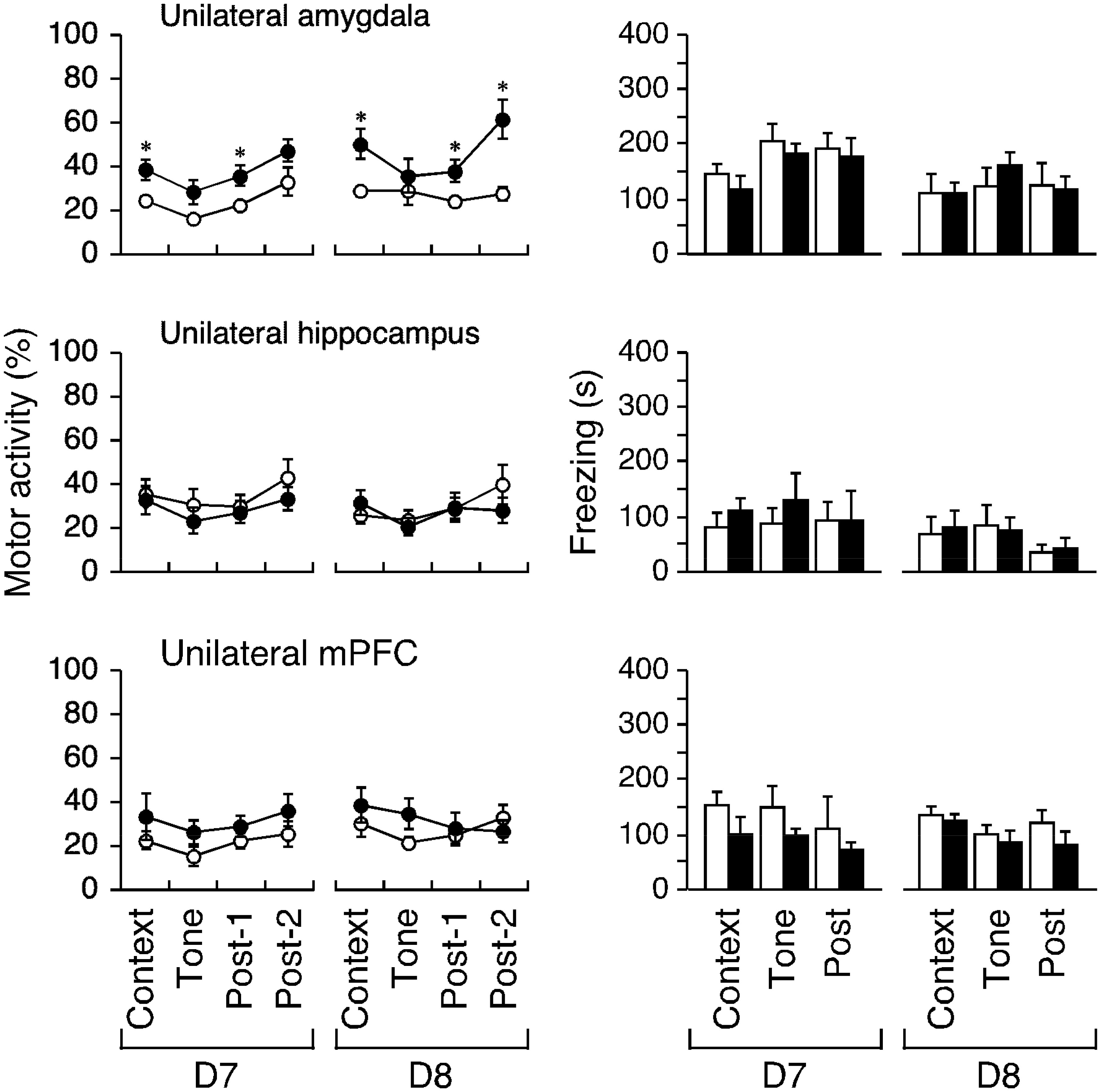
Figures(7)

Masatoshi Takita, Yumi Izawa-Sugaya. Neurocircuit differences between memory traces of persistent hypoactivity and freezing following fear conditioning among the amygdala, hippocampus, and prefrontal cortex[J]. AIMS Neuroscience, 2021, 8(2): 195-211. doi: 10.3934/Neuroscience.2021010







 DownLoad:
DownLoad: