Citation: Natalie Ceballos, Shobhit Sharma. Risk and Resilience: The Role of Brain-derived Neurotrophic Factor in Alcohol Use Disorder[J]. AIMS Neuroscience, 2016, 3(4): 398-432. doi: 10.3934/Neuroscience.2016.4.398

| [1] |
Logrip ML, Segev B, Warnault V, et al. (2015) Corticostriatal BDNF and alcohol addiction. Brain Res 1628: 60-67. doi: 10.1016/j.brainres.2015.03.025

|
| [2] | World Health Organization (2014) Global status report on alcohol and health. Available from: http://www.who.int/substance_abuse/publications/global_alcohol_report/en/ |
| [3] |
Cadet JL (2016) Epigenetics of stress, addiction and resilience: therapeutic implications. Mol Neurobiol 53: 545-560. doi: 10.1007/s12035-014-9040-y
|
| [4] |
Martinez JM, Garakani A, Yehuda R et al. (2012) Proinflammatory and “resiliency” proteins in the CSF of patients with major depression. Depress Anxiety 29: 32-38. doi: 10.1002/da.20876
|
| [5] | Daskalakis NP, de Kloet E, Yehuda R, et al. (2015) Early life stress effects on glucocorticoid-BDNF interplay in the hippocampus. Front Mol Neurosci 8: 68-80. |
| [6] |
Pang PT, Teng HK, Zaitsev E, et al. (2004) Cleavage of proBDNF by tPA/plasmin is essential for long-term hippocampal plasticity. Science 306: 487-491. doi: 10.1126/science.1100135
|
| [7] |
Revest JM, Le Roux A, Roullot-Lacarriere V, et al. (2014) BDNF-TrkB signaling through Erk1/2 MAPK phosphorylation mediates the enhancement of fear memory induced by glucocorticoids. Mol Psychiatry 19: 1001-1009. doi: 10.1038/mp.2013.134
|
| [8] |
Chao MV (2003) Neurotrophins and their receptors: a convergence point for many signaling pathways. Nat Rev Neurosci 4: 299-309. doi: 10.1038/nrn1078
|
| [9] |
Huang EJ, Reichardt LF (2003) Trk receptors: roles in neuronal signal transduction. Ann Rev Biochem 72: 609-642. doi: 10.1146/annurev.biochem.72.121801.161629
|
| [10] | Egan MF, Kojima M, Callicott JH (2003) The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell 112: 257-269. |
| [11] |
Lu B, Gottschalk W (2000) Modulation of hippocampal synaptic transmission and plasticity by neurotrophins. Prog Brain Res 128: 231-241. doi: 10.1016/S0079-6123(00)28020-5
|
| [12] |
Poo MM (2001) Neurotrophins as synaptic modulators. Nat Rev Neurosci 2: 24-32. doi: 10.1038/35049004
|
| [13] | Koshimizu H, Kiyosue K, Hara T, et al. (2009) Multiple functions of precursor BDNF to CNS neurons: negative regulation of neurite growth, spine formation and cell survival. Mol Brain 13: 2-27. |
| [14] |
Hosang GM, Shiles C, Tansey KE, et al. (2014) Interaction between stress and the BDNF Val66Met polymorphism in depression: a systematic review and meta-analysis. BMC Med 12: 7-18. doi: 10.1186/1741-7015-12-7
|
| [15] |
Chen ZY, Patel PD, Sant G, et al. (2004) Variant brain-derived neurotrophic factor (BDNF) (Met 66) alters the intracellular trafficking and activity-dependent secretion of wild-type BDNF in neurosecretory cells and cortical neurons. J Neurosci 24: 4401-4411. doi: 10.1523/JNEUROSCI.0348-04.2004
|
| [16] |
Czira ME, Wersching H, Baune BT, et al. (2012) Brain-derived neurotrophic factor gene polymorphisms, neurotransmitter levels, and depressive symptoms in an elderly population. Age (Dordr) 34: 1529-1541. doi: 10.1007/s11357-011-9313-6
|
| [17] |
Hong CJ, Liou YJ, Tsai SJ (2011) Effects of BDNF polymorphisms on brain function and behavior in health and disease. Brain Res Bull 86: 287-297. doi: 10.1016/j.brainresbull.2011.08.019
|
| [18] |
Cheeran B, Talelli P, Mori F, et al. (2008) A common polymorphism in the brain-derived neurotrophic factor gene (BDNF) modulates human cortical plasticity and response to rTMS. J Physiol 586: 5717-5725. doi: 10.1113/jphysiol.2008.159905
|
| [19] | Lamb YN, McKay NS, Thompson CS, et al. (2015a) Brain-derived neurotrophic factor Val66Met polymorphism, human memory, and synaptic neuroplasticity. Wiley Interdiscip Rev Cogn Sci 6: 97-108. |
| [20] | Lamb YN, Thompson CS, McKay NS, et al. (2015b) The brain-derived neurotrophic factor (BDNF) val66met polymorphism differentially affects performance on subscales of the Wechsler Memory Scale—Third Edition (WMS-III). Front Psychol 6: 1212-1220. |
| [21] | Hariri AR, Goldberg TE, Mattay VS, et al. (2003) Brain-derived neurotrophic factor val66met polymorphism affects human memory-related hippocampal activity and predicts memory performance. J Neurosci 23: 6690-6694. |
| [22] | Notaras M, Hill R, van den Buuse M (2015a) The BDNF gene Val66Met polymorphism as a modifier of psychiatric disorder susceptibility: progress and controversy. Mol Psychiatry 20: 916-930. |
| [23] | Notaras M, Hill R, van den Buuse M (2015b) A role for the BDNF gene Val66Met polymorphism in schizophrenia? A comprehensive review. Neurosci Biobehav Rev 51: 15-30. |
| [24] | Feltenstein MW, See RE (2013) Systems level neuroplasticity in drug addiction. Cold Spring Harb Perspect Med 3: a011916 |
| [25] |
Luscher C, Malenka RC (2011) Drug-evoked synaptic plasticity in addiction: from molecular changes to circuit remodeling. Neuron 69: 650-663. doi: 10.1016/j.neuron.2011.01.017
|
| [26] |
Nestler EJ (2014) Epigenetic mechanisms of drug addiction. Neuropharmacology 76: 259-268. doi: 10.1016/j.neuropharm.2013.04.004
|
| [27] | Russo SJ, Dietz DM, Dumitriu D, et al. (2011) The addicted synapse: mechanisms of synaptic and structural plasticity in the nucleus accumbens. Trends Neurosci 33: 267-276. |
| [28] | Moonat S, Pandey SC (2012) Stress, epigenetics, and alcoholism. Alcohol Res 34: 495-505. |
| [29] |
Ghitza UE, Zhai H, Wu P, et al. (2010) Role of BDNF and GDNF in drug reward and relapse: a review. Neurosci Biobehav Rev 35: 157-171. doi: 10.1016/j.neubiorev.2009.11.009
|
| [30] |
Logrip ML, Janak PH, Ron D (2009) Escalating ethanol intake is associated with altered corticostriatal BDNF expression. J Neurochem 109: 1459-1468. doi: 10.1111/j.1471-4159.2009.06073.x
|
| [31] | Darcq E, Warnault V, Phamluong K, et al. (2015) MicroRNA-30a-5p in the prefrontal cortex controls the transition from moderate to excessive alcohol consumption. Mol Psychiatry 20: 1219-1231. |
| [32] |
Jeanblanc J, He D-Y, Carnicella S, et al. (2009) Endogenous BDNF in the dorsolateral striatum gates alcohol drinking. J Neurosci 29: 13494-13502. doi: 10.1523/JNEUROSCI.2243-09.2009
|
| [33] |
McGough NNH, He D-Y, Logrip ML, et al. (2004) RACK1 and brain-derived neurotrophic factor: a homeostatic pathway that regulates alcohol addiction. J Neurosci 24: 10542-10552. doi: 10.1523/JNEUROSCI.3714-04.2004
|
| [34] |
Logrip ML, Janak PH, Ron D (2008) Dynorphin is a downstream effector of striatal BDNF regulation of ethanol intake. FASEB Journal 22: 2393-2404. doi: 10.1096/fj.07-099135
|
| [35] | Vengeliene V, Bilbao A, Molander A, et al. (2008) Neuropharmacology of alcohol addiction. Br J Pharmacol 154: 299-315. |
| [36] |
Spanagel R (2009) Alcoholism: a systems approach from molecular physiology to addictive behavior. Physiol Rev 89: 649-705. doi: 10.1152/physrev.00013.2008
|
| [37] |
Breese GR, Criswell HE, Carta M, et al. (2006) Basis of the gabamimetic profile of ethanol. Alcohol Clin Exp Res 30: 731-744. doi: 10.1111/j.0145-6008.2006.00086.x
|
| [38] |
Roberto M, Madamba SG, Moore SD, et al. (2003). Ethanol increases GABAergic transmission at both pre- and postsynaptic sites in rat central amygdala neurons. Proc Natl Acad Sc. USA 100: 2053-2058. doi: 10.1073/pnas.0437926100
|
| [39] |
Silberman Y, Winder DG (2015) Ethanol and corticotropin releasing factor receptor modulation of central amygdala neurocircuitry: an update and future directions. Alcohol 49: 179-184. doi: 10.1016/j.alcohol.2015.01.006
|
| [40] |
Gilpin NW, Herman MA, Roberto M (2015) The central amygdala as an integrative hub for anxiety and alcohol use disorders. Biol Psychiatry 77: 859-869. doi: 10.1016/j.biopsych.2014.09.008
|
| [41] | Brady KT, Sonne SC (1999) The role of stress in alcohol use, alcoholism treatment, and relapse. Alcohol Res Health 23: 263-271. |
| [42] |
Sinha R (2001) How does stress increase risk of drug abuse and relapse? Psychopharmacology 158: 343-359. doi: 10.1007/s002130100917
|
| [43] |
Sinha R (2008) Chronic stress, drug use, and vulnerability to addiction. Ann N Y Acad Sci 1141: 105-130. doi: 10.1196/annals.1441.030
|
| [44] | Becker HC (2012) Effects of alcohol dependence and withdrawal on stress responsiveness and alcohol consumption. Alc Res 34: 448-458. |
| [45] |
Koob G, Kreek MJ (2007) Stress, dysregulation of drug reward pathways, and the transition to drug dependence. Am J Psychiatry 164: 1149-1159. doi: 10.1176/appi.ajp.2007.05030503
|
| [46] |
Koob GF, Le Moal M (1997) Drug abuse: hedonic homeostatic dysregulation. Science 278: 52-58. doi: 10.1126/science.278.5335.52
|
| [47] | Koob GF, Volkow ND (2010) Neurocircuitry of addiction. Neuropsychopharmacology 35: 217-238. |
| [48] |
Vink JM (2016) Genetics of addiction: future focus on gene x environment interaction? J Stud Alcohol Drugs 77: 684-687. doi: 10.15288/jsad.2016.77.684
|
| [49] |
Morrow JD, Flagel SB (2016) Neuroscience of resilience and vulnerability for addiction medicine: from genes to behavior. Prog Brain Res 223: 3-18. doi: 10.1016/bs.pbr.2015.09.004
|
| [50] |
Pittenger C (2011) Epigenetic modification of the BDNF locus by early-life enrichment: towards a molecular correlate of resilience? Neurosci Lett 495: 165-167. doi: 10.1016/j.neulet.2011.03.041
|
| [51] |
Cecil CAM, Walton E, Viding E (2016) Epigenetics of alcoholism: current knowledge, challenges, and future directions. J Stud Alcohol Drugs 77: 688-691. doi: 10.15288/jsad.2016.77.688
|
| [52] |
Malter Cohen M, Tottenham N, Casey BJ (2013) Translational developmental studies of stress on brain and behavior: implications for adolescent mental health and illness? Neuroscience 249: 53-62. doi: 10.1016/j.neuroscience.2013.01.023
|
| [53] |
Jeanneteau F, Chao MV (2013) Are BDNF and glucocorticoid activities calibrated? Neuroscience 239: 173-195. doi: 10.1016/j.neuroscience.2012.09.017
|
| [54] |
Jeanneteau FD, Lambert SM, Ismaili N, et al. (2012) BDNF and glucocorticoids regulate corticotrophin-releasing hormone (CRH) homeostasis in the hypothalamus. Proc Natl Acad Sci USA 109: 1305-1310. doi: 10.1073/pnas.1114122109
|
| [55] |
McEwen BS, Bowles NP, Gray JD, et al. (2015) Mechanisms of stress in the brain. Nat Neurosci 18: 1353-1363. doi: 10.1038/nn.4086
|
| [56] | de Kloet E, Joels M, Holsboer F (2005) Stress and the brain: from adaptation to disease. Nat Rev Neurosci 6: 463-475. |
| [57] |
Davidson RJ, McEwen BS (2012) Social influences on neuroplasticity: stress and interventions to promote well-being. Nat Neurosci 15: 689-695. doi: 10.1038/nn.3093
|
| [58] |
Grassi-Oliveira R, Stein LM, Lopes RP, et al. (2008) Low plasma brain-derived neurotrophic factor and childhood physical neglect are associated with verbal memory impairment in major depression – a preliminary report. Biol Psychiatry 64: 281-285. doi: 10.1016/j.biopsych.2008.02.023
|
| [59] |
Roth TL, Lubin FD, Funk AJ, et al. (2009) Lasting epigenetic influence of early-life adversity on the BDNF gene. Biol Psychiatry 65: 760-769. doi: 10.1016/j.biopsych.2008.11.028
|
| [60] |
Branchi I, Karpova NN, D’Andrea I, et al. (2011) Epigenetic modifications induced by early enrichment are associated with changes in timing of induction of BDNF expression. Neurosci Lett 495: 168-172. doi: 10.1016/j.neulet.2011.03.038
|
| [61] |
Forero DA, Lopez-Leon S, Shin HD, et al. (2015) Meta-analysis of six genes (BDNF, DRD1, DRD3, GRIN2B and MAOA) involved in neuroplasticity and the risk for alcohol dependence. Drug Alcohol Depend 149: 259-263. doi: 10.1016/j.drugalcdep.2015.01.017
|
| [62] | Haerian BS (2013) BDNF rs6265 polymorphism and drug addiction: a systematic review and meta-analysis. Pharmacogenomics 14: 2055-2065. |
| [63] |
Meng D, Wu T, Rao U, et al. (2011) Serum NPY and BDNF response to a behavioral stressor in alcohol-dependent and healthy control participants. Psychopharmacology 218: 59-67. doi: 10.1007/s00213-011-2414-1
|
| [64] |
Karege F, Bondolfi G, Gervasoni N, et al. (2005) Low brain-derived neurotrophic factor (BDNF) levels in serum of depressed patients probably results from lowered platelet BDNF release unrelated to platelet reactivity. Biol Psychiatry 57: 1068-1072. doi: 10.1016/j.biopsych.2005.01.008
|
| [65] | Unternaehrer E, Luers P, Mill J, et al. (2012). Dynamic changes in DNA methylation of stress-associated genes (OXTR, BDNF) after acute psychosocial stress. Transl Psychiatry 2: e150 |
| [66] |
Giese M, Beck J, Brand S (2014) Fast BDNF serum level increase and diurnal BDNF oscillations are associated with therapeutic response after partial sleep deprivation. J Psychiatr Res 59: 1-7. doi: 10.1016/j.jpsychires.2014.09.005
|
| [67] |
Pan W, Banks WA, Fasold MB, et al. (1998) Transport of brain-derived neurotrophic factor across the blood-brain barrier. Neuropharmacology 37: 1553-1561. doi: 10.1016/S0028-3908(98)00141-5
|
| [68] |
D’Sa C, Dileone RJ, Anderson GM, et al. (2012) Serum and plasma brain-derived neurotrophic factor (BDNF) in abstinent alcoholics and social drinkers. Alcohol 46: 253-259. doi: 10.1016/j.alcohol.2011.12.001
|
| [69] | Klein AB, Williamson R, Santini MA, et al. (2010) Blood BDNF concentrations reflect brain-tissue BDNF levels across species. Int J Neuropsychopharmacol 7: 1-7. |
| [70] |
Sartorius A, Hellweg R, Litzke J, et al. (2009) Correlations and discrepancies between serum and brain tissue levels of neurotrophins after electroconvulsive treatment in rats. Pharmacopsychiatry 42: 270-276. doi: 10.1055/s-0029-1224162
|
| [71] |
Lang UE, Hellweg R, Seifert F, et al. (2007) Correlation between serum brain-derived neurotrophic factor level and an in vivo marker of cortical integrity. Biol Psychiatry 62: 530-535. doi: 10.1016/j.biopsych.2007.01.002
|
| [72] |
Bus BAA, Molendijk ML, Penninx BJWH, et al. (2011) Determinants of serum brain-derived neurotrophic factor. Psychoneuroendocrinology 36: 228-239. doi: 10.1016/j.psyneuen.2010.07.013
|
| [73] |
Begliuomini E, Lenzi F, Ninni E, et al. (2008) Plasma brain-derived neurotrophic factor daily variations in men: correlation with cortisol circadian rhythm. J Endocrinol 197: 429-435. doi: 10.1677/JOE-07-0376
|
| [74] |
Pluchino N, Cubeddu A, Begliuomini S, et al. (2009) Daily variation of brain-derived neurotrophic factor and cortisol in women with normal menstrual cycles, undergoing oral contraception and in postmenopause. Hum Reprod 24: 2303-2309. doi: 10.1093/humrep/dep119
|
| [75] |
Choi S-W, Bhang S, Ahn J-H (2011) Diurnal variation and gender differences of plasma brain-derived neurotrophic factor in healthy human subjects. Psychiatry Res 186: 427-430. doi: 10.1016/j.psychres.2010.07.028
|
| [76] | Oh H, Lewis DA, Sibille E (2016) The role of BDNF in age-dependent changes of excitatory and inhibitory synaptic markers in the human prefrontal cortex. Neuropsychopharmacology [Epub ahead of print, doi: 10.1038/npp.2016.126]. |
| [77] | Del Boca F, Darkes J (2003) The validity of self-reports of alcohol consumption: state of the science and challenges for research. Addiction 98: 1-12. |
| [78] | Sobell LC, Sobell MB (1995) Alcohol consumption measures. In: Allen, J.P., Columbus, M. Editors, Assessing Alcohol Problems: A Guide for Clinicians and Researchers, Bethesda, MD: National Institute on Alcohol Abuse & Alcoholism, 55-73. |
| [79] | Leeman RF, Heilig M, Cunningham CL, et al. (2010) Ethanol consumption: how should we measure it? Achieving consilience between human and animal phenotypes. Addict Biol 15: 109-124. |
| [80] | Thabrew H, de Sylva S, Romans SE (2012) Evaluating childhood adversity. Adv Psychosom Med 32: 35-57. |
| [81] |
Brown GW, Craig TK, Harris TO, et al. (2007) Validity of retrospective measures of early maltreatment and depressive episodes using the childhood experience of care and abuse (ceca) instrument—a life-course study of adult chronic depression-2. J Affect Disord 103: 217-224. doi: 10.1016/j.jad.2007.06.003
|
| [82] |
Fergusson DM, Horwood LJ, Woodward LJ (2000) The stability of child abuse reports: a longitudinal study of the reporting behavior of young adults. Psychol Med 30: 529-544. doi: 10.1017/S0033291799002111
|
| [83] |
Shin S, Stewart R, Ferri CP, et al. (2010) An investigation of associations between alcohol use disorder and polymorphisms on ALDH2, BDNF, 5-HTTLPR, and MTHFR genes in older Korean men. Int J Geriatr Psychiatry 25: 441-448. doi: 10.1002/gps.2358
|
| [84] |
Colzato LS, Van der Does AJ, Kouwenhoven C, et al. (2011) BDNF Val66Met polymorphism is associated with higher anticipatory cortisol stress response, anxiety, and alcohol consumption in healthy adults. Psychoneuroendocrinology 36: 1562-1569. doi: 10.1016/j.psyneuen.2011.04.010
|
| [85] |
Su N, Zhang L, Fei F, et al. (2011) The brain-derived neurotrophic factor is associated with alcohol dependence-related depression and antidepressant response. Brain Res 1415: 119-126. doi: 10.1016/j.brainres.2011.08.005
|
| [86] |
Cheah SY, Lawford BR, Young RM, et al. (2014) BDNF SNPs are implicated in comorbid alcohol dependence in schizophrenia but not in alcohol-dependent patients without schizophrenia. Alcohol Alcohol 49: 491-497. doi: 10.1093/alcalc/agu040
|
| [87] | Nees F, Witt SH, Dinu-Biringer R, et al. (2015) BDNF Val66Met and reward-related brain function in adolescents: role for early alcohol consumption. Alcohol 49: 103-110. |
| [88] |
Hill SY, Wang S, Carter H, et al. (2011) Cerebellum volume in high-risk offspring from multiplex alcohol dependence families: association with allelic variation in GABRA2 and BDNF. Psychiatry Res 194: 304-313. doi: 10.1016/j.pscychresns.2011.05.006
|
| [89] |
Kim SG, Ugurbil K, Strick PL (1994) Activation of a cerebellar output nucleus during cognitive processing. Science 265: 949-951. doi: 10.1126/science.8052851
|
| [90] |
Andreasen NC, O’Leary DS, Cizaldo T, et al. (1995) PET studies of memory: novel versus practiced free recall of word lists. Neuroimage 2: 296-305. doi: 10.1006/nimg.1995.1037
|
| [91] |
Andreasen NC, O’Leary DS, Arndt S, et al. (1996) Neural substrates of facial recognition. J Neuropsychiatry Clin Neurosci 8: 139-146. doi: 10.1176/jnp.8.2.139
|
| [92] | Andreasen NC, O’Leary DS, Paradiso S, et al. (1999). The cerebellum plays a role in conscious episodic memory retrieval. Hum Brain Mapp 8: 226-234. |
| [93] |
Allen G, Buxton R, Wong E, et al. (1997) Attentional activation of the cerebellum independent of motor involvement. Science 275: 1940-1943. doi: 10.1126/science.275.5308.1940
|
| [94] | Fiez JA, Raichle ME, Miezin FM, et al. (1995) PET studies of auditory and phonological processing: effects of stimulus characteristics and task demands. Trends Cogn Sci 7: 357-375. |
| [95] |
Turner BM, Paradiso S, Marvel C, et al. (2007) The cerebellum and emotional experience. Neuropsychologia 45: 1331-1341. doi: 10.1016/j.neuropsychologia.2006.09.023
|
| [96] |
Mon A, Durazzo TC, Gazdzinski S, et al. (2013) Brain-derived neurotrophic factor genotype is associated with brain gray and white matter tissue volumes recovery in abstinent alcohol-dependent individuals. Genes Brain Behav 12: 98-107. doi: 10.1111/j.1601-183X.2012.00854.x
|
| [97] |
Hoefer ME, Pennington DL, Durazzo TC, et al. (2014) Genetic and behavioral determinants of hippocampal volume recovery during abstinence from alcohol. Alcohol 48: 631-638. doi: 10.1016/j.alcohol.2014.08.007
|
| [98] | Pecina M, Martinez-Jauand M, Love T, et al. (2014) Valence-specific effects of BDNF Val66Met polymorphism on dopaminergic stress and reward processing in humans. J Neurosci 34: 5874-5881. |
| [99] |
Chen J, Hutchison KE, Calhoun VD, et al. (2015) CREB-BDNF pathway influences alcohol cue-elicited activation in drinkers. Hum Brain Mapp 36: 3007-3019. doi: 10.1002/hbm.22824
|
| [100] | Claus ED, Ewing SW, Filbey FM, et al. (2011) Identifying neurobiological phenotypes associated with alcohol use disorder severity. Neuropsychopharmacology 36: 2086-2096. |
| [101] | Liu J, Calhoun VD, Chen J, et al. (2013) Effect of homozygous deletions at 22q13.1 on alcohol dependence severity and cue-elicited BOLD response in the precuneus. Addict Biol 18: 548-558. |
| [102] |
Park MS, Sohn JH, Suk JA, et al. (2007) Brain substrates of craving to alcohol cues in subjects with alcohol use disorder. Alcohol Alcohol 42: 417-422. doi: 10.1093/alcalc/agl117
|
| [103] |
Schacht JP, Anton RF, Myrick H. (2013) Functional neuroimaging studies of alcohol cue reactivity: a quantitative meta-analysis and systematic review. Addict Biol 18: 121-133. doi: 10.1111/j.1369-1600.2012.00464.x
|
| [104] |
Tapert SF, Brown GG, Baratta MV, et al. (2004) fMRI BOLD response to alcohol stimuli in alcohol dependent young women. Addict Behav 29: 33-50. doi: 10.1016/j.addbeh.2003.07.003
|
| [105] |
Grzywacz A, Samochowiec A, Ciechanowicz A, et al. (2010) Family-based study of brain-derived neurotrophic factor (BDNF) gene polymorphism in alcohol dependence. Pharmacol Rep 62: 938-941. doi: 10.1016/S1734-1140(10)70354-6
|
| [106] |
Muschler MA, Heberlein A, Frieling H, et al. (2011) Brain-derived neurotrophic factor, Val66Met single nucleotide polymorphism is not associated with alcohol dependence. Psychiatr Genet 21: 53-54. doi: 10.1097/YPG.0b013e32834133ab
|
| [107] |
Benzerouk F, Gierski F, Gorwood P, et al. (2013) Brain-derived neurotrophic factor (BDNF) Val66Met polymorphism and its implication in executive functions in adult offspring of alcohol-dependent probands. Alcohol 47: 271-274. doi: 10.1016/j.alcohol.2013.03.001
|
| [108] |
Benzerouk F, Gierski F, Raucher-Chene D, et al. (2015) Association study between reward dependence and a functional BDNF polymorphism in adult women offspring of alcohol-dependent probands. Psychiatr Genet 25: 208-211. doi: 10.1097/YPG.0000000000000099
|
| [109] |
Nedic G, Perkovic MN, Sviglin KN, et al. (2013) Brain-derived neurotrophic factor Val66Met polymorphism and alcohol-related phenotypes. Prog Neuropsychopharmacol Biol Psychiatry 40: 193-198. doi: 10.1016/j.pnpbp.2012.09.005
|
| [110] | Serretti A, Souery D, Antypa N, et al. (2013) The impact of adverse life events on clinical features and interaction with gene variants in mood disorder patients. Psychopathology 46: 384-389. |
| [111] | Dalvie S, Stein DJ, Koenen K, et al. (2014) The BDNF p.Val66Met polymorphism, childhood trauma, and brain volumes in adolescents with alcohol abuse. BMC Psychiatry 14: 328-335. |
| [112] |
Heberlein A, Buscher P, Schuster R, et al. (2015) Do changes in the BDNF promotor methylation indicate the risk of alcohol relapse? Eur Neuropsychopharmacol 25: 1892-1897. doi: 10.1016/j.euroneuro.2015.08.018
|
| [113] | Moore D, (2015) The Developing Genome. Oxford: Oxford University Press. |
| [114] | Morrow JD, Flagel SB (2016) Neuroscience of resilience and vulnerability for addiction medicine: from genes to behavior. |
| [115] |
Lhullier AC, Moreira FP, da Silva RA, et al. (2015) Increased serum neurotrophin levels related to alcohol use disorder in a young population sample. Alcohol Clin Exp Res 39: 30-33. doi: 10.1111/acer.12592
|
| [116] |
Zhang XY, Tan YL, Chen DC, et al. (2016) Effects of cigarette smoking and alcohol use on neurocognition and BDNF levels in a Chinese population. Psychopharmacology 233: 435-445. doi: 10.1007/s00213-015-4124-6
|
| [117] |
Buchsbaum DG, Buchanan RG, Welsh J, et al. (1992) Screening for drinking disorders in the elderly using the CAGE questionnaire. J Am Geriatr Soc 40: 662-665. doi: 10.1111/j.1532-5415.1992.tb01956.x
|
| [118] |
Selzer ML (1971) The Michigan alcoholism screening test: the quest for a new diagnostic instrument. Am J Psychiatry 127: 1653-1658. doi: 10.1176/ajp.127.12.1653
|
| [119] |
Heberlein A, Muschler M, Wilhelm J, et al. (2010) BDNF and GDNF serum levels in alcohol-dependent patients during withdrawal. Prog Neuropsychopharmacol Biol Psychiatry 34: 1060-1064. doi: 10.1016/j.pnpbp.2010.05.025
|
| [120] |
Costa MA, Girard M, Dalmay F, et al. (2011) Brain-derived neurotrophic factor serum levels in alcohol-dependent subjects 6 months after alcohol withdrawal. Alcohol Clin Exp Res 35: 1966-1973. doi: 10.1111/j.1530-0277.2011.01548.x
|
| [121] | Zanardini R, Fontana A, Pagano R, et al. (2011) Alterations of brain-derived neurotrophic factor serum levels in patients with alcohol dependence. Alcohol Clin Exp Res 35: 1529-1533. |
| [122] |
Huang MC, Chen CH, Liu HC, et al. (2011) Differential patterns of serum brain-derived neurotrophic factor levels in alcoholic patients with and without delirium tremens during acute withdrawal. Alcohol Clin Exp Res 35: 126-131. doi: 10.1111/j.1530-0277.2010.01329.x
|
| [123] |
Kohler S, Klimke S, Hellweg R, et al. (2013) Serum brain-derived neurotrophic factor and nerve growth factor concentrations change after alcohol withdrawal: preliminary data of a case-control comparison. Eur Addict Res 19: 98-104. doi: 10.1159/000342334
|
| [124] |
Reynolds PM, Mueller SW, MacLaren R, (2015) A comparison of dexmedetomidine and placebo on the plasma concentrations of NGF, BDNF, GDNF, and epinephrine during severe alcohol withdrawal. Alcohol 49: 15-19. doi: 10.1016/j.alcohol.2014.11.006
|
| [125] | Kirschbaum C, Pirke KM, Hellhammer DH. (1993) The ‘Trier Social Stress Test’—a tool for investigating psychobiological stress responses in a laboratory setting. Neuropsychobiology 28: 76-81. |
| [126] | Kim JH, Kim SJ, Lee WY, et al. (2013) The effects of alcohol abstinence on BDNF, ghrelin, and leptin secretions in alcohol-dependent patients with glucose tolerance. Alcohol Clin Exp Res 37 Suppl 1: E52-E58. |
| [127] | Miguez-Burbano MJ, Espinoza L, Whitehead NE, et al. (2014a) Brain-derived neurotrophic factor and cognitive status: the delicate balance among people living with HIV, with and without alcohol abuse. Curr HIV Res 12: 254-264. |
| [128] | Miguez-Burbano MJ, Espinoza L, Vargas et al. (2014b) Mood disorders and BDNF relationship with alcohol drinking trajectories among PLWH receiving care. J Alcohol Drug Depend 2: 148-165. |
| [129] | Skinner HA, Horn JL, (1984) Alcohol Dependence Scale (ADS) user’s guide. Toronto: Addiction Research Foundation. |
| [130] | Parsons OA, Nixon SJ (1993) Neurobehavioral sequelae of alcoholism. Neurol Clin 11: 205-218. |
| [131] | Neupane SP, Lien L, Ueland T, et al. (2015) Serum brain-derived neurotrophic factor levels in relation to comorbid depression and cytokine levels in Nepalese men with alcohol-use disorders. Alcohol 49: 471-478. |
| [132] |
Heath AC, Lessov-Schlaggar CN, Lian M, et al. (2016) Research on gene-environment interplay in the era of “big data”. J Stud Alcohol Drugs 77: 681-683. doi: 10.15288/jsad.2016.77.681
|
| [133] |
Dick DM (2016) The genetics of addiction: where do we go from here? J Stud Alcohol Drugs 77: 673-675. doi: 10.15288/jsad.2016.77.673
|
| [134] | Duncan, L.E., & Keller, M.C. (2011). A critical review of the first 10 years of candidate gene-by-environment interaction research in psychiatry. Am J Psychiatry 168: 1041-1049. |
| [135] | Srinivasan S, Shariff M, Bartlett SE (2013) The role of the glucocorticoids in developing resilience to stress and addiction. Front Psychiatry 4: 1-11. |
| [136] |
Erickson KI, Voss MW, Prakash RS, et al. (2011) Exercise training increases size of hippocampus and improves memory. Proc Natl Acad Sci USA 108: 3017-3022. doi: 10.1073/pnas.1015950108
|
| [137] |
Disner SG, Beevers CG, Haigh EA, et al. (2011) Neural mechanisms of the cognitive model of depression. Nat Rev Neurosci 12: 467-477. doi: 10.1038/nrn3027
|
| [138] |
Clark DA, Beck AT (2010) Cognitive theory and therapy of anxiety and depression: convergence with neurobiological findings. Trends Cogn Sci 14: 418-424. doi: 10.1016/j.tics.2010.06.007
|
| [139] |
Lutz A, Slagter HA, Dunne JD, et al. (2008) Attention regulation and monitoring in meditation. Trends Cogn Sci 12: 163-169. doi: 10.1016/j.tics.2008.01.005
|
| [140] |
Morris T (2016) Developing the catecholamines hypothesis for the acute exercise-cognition interaction in humans: lessons from animal studies. Physiol Behav 165: 291-299. doi: 10.1016/j.physbeh.2016.08.011
|
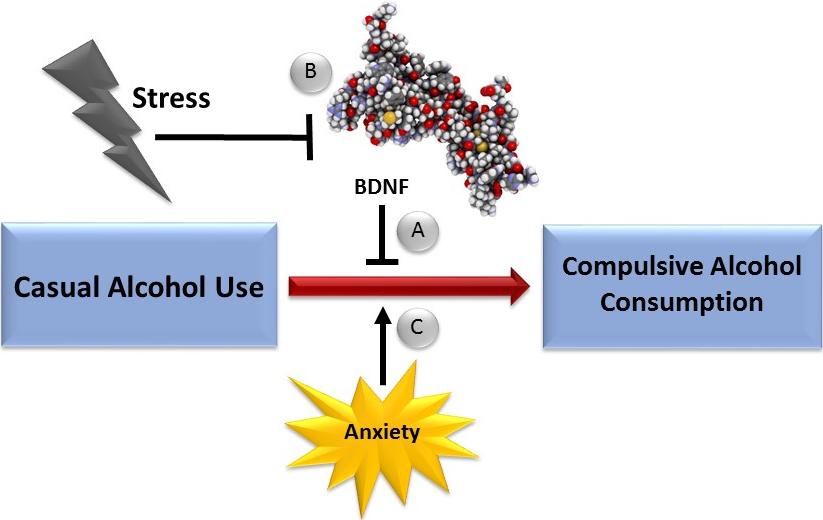
Figures(1) / Tables(2)

Natalie Ceballos, Shobhit Sharma. Risk and Resilience: The Role of Brain-derived Neurotrophic Factor in Alcohol Use Disorder[J]. AIMS Neuroscience, 2016, 3(4): 398-432. doi: 10.3934/Neuroscience.2016.4.398







 DownLoad:
DownLoad: