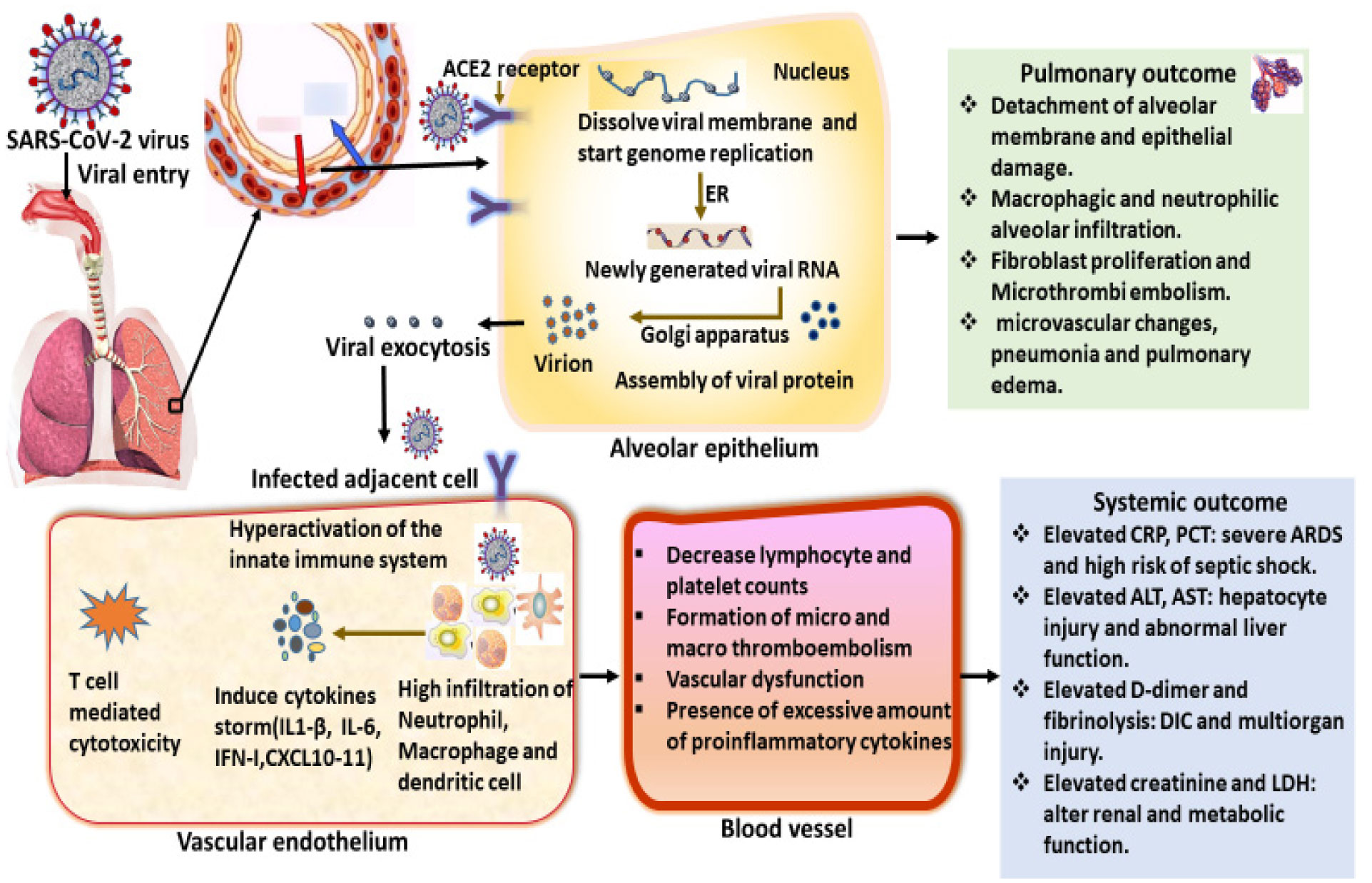
Coronavirus disease 2019 (COVID-19) is highly infectious and may induce epigenetic alteration of the host immune system. Understanding the role of epigenetic mechanisms in COVID-19 infection is a clinical need to minimize critical illness and widespread transmission. The susceptibility to infection and progression of COVID-19 varies from person to person; pathophysiology substantially depends on epigenetic changes in the immune system and preexisting health conditions. Recent experimental and epidemiological studies have revealed the method of transmission and clinical presentation related to COVID-19 pathogenesis, however, the underlying pathology of variation in the severity of infection remains questionable. Epigenetic changes may also be responsible factors for multisystem association and deadly systemic complications of severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) infected patients. Commonly, epigenetic changes are evoked by alteration of the host's immune response, stress, preexisting condition, oxidative stress response, external behavioral or environmental factors, and age. In addition, the viral infection itself might manipulate the host immune responses associated with inflammation by reprogramming epigenetic processes which are the susceptible factor for disease severity and death. As a result, epigenetic events such as histone modification and DNA methylation are implicated in regulating pro-inflammatory cytokines production by remodeling macrophage and T-cell activity towards inflammation, consequently, may also affect tissue repair and injury resolution process. This review aims to discuss the comprehensive understanding of the epigenetic landscape of COVID-19 disease progression that varies from person to person with supporting interdisciplinary prognosis protocol to overcome systemic impairment.
Citation: Zinia Pervin, Anika Tasnim, Hasib Ahamed, Md Al Hasibuzzaman. Epigenetic regulation of the COVID-19 pathogenesis: its impact on the host immune response and disease progression[J]. AIMS Allergy and Immunology, 2023, 7(1): 60-81. doi: 10.3934/Allergy.2023005
Coronavirus disease 2019 (COVID-19) is highly infectious and may induce epigenetic alteration of the host immune system. Understanding the role of epigenetic mechanisms in COVID-19 infection is a clinical need to minimize critical illness and widespread transmission. The susceptibility to infection and progression of COVID-19 varies from person to person; pathophysiology substantially depends on epigenetic changes in the immune system and preexisting health conditions. Recent experimental and epidemiological studies have revealed the method of transmission and clinical presentation related to COVID-19 pathogenesis, however, the underlying pathology of variation in the severity of infection remains questionable. Epigenetic changes may also be responsible factors for multisystem association and deadly systemic complications of severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) infected patients. Commonly, epigenetic changes are evoked by alteration of the host's immune response, stress, preexisting condition, oxidative stress response, external behavioral or environmental factors, and age. In addition, the viral infection itself might manipulate the host immune responses associated with inflammation by reprogramming epigenetic processes which are the susceptible factor for disease severity and death. As a result, epigenetic events such as histone modification and DNA methylation are implicated in regulating pro-inflammatory cytokines production by remodeling macrophage and T-cell activity towards inflammation, consequently, may also affect tissue repair and injury resolution process. This review aims to discuss the comprehensive understanding of the epigenetic landscape of COVID-19 disease progression that varies from person to person with supporting interdisciplinary prognosis protocol to overcome systemic impairment.

| [1] |
Kahn JS, McIntosh K (2005) History and recent advances in coronavirus discovery. Pediatr Infect Dis J 24: S223-S227. https://doi.org/10.1097/01.inf.0000188166.17324.60 
|
| [2] |
Isaacs D, Flowers D, Clarke JR, et al. (1983) Epidemiology of coronavirus respiratory infections. Arch Dis Child 58: 500-503. https://doi.org/10.1136/adc.58.7.500
|
| [3] |
Weiss SR, Navas-Martin S (2005) Coronavirus pathogenesis and the emerging pathogen severe acute respiratory syndrome coronavirus. Microbiol Mol Biol R 69: 635-664. https://doi.org/10.1128/MMBR.69.4.635-664.2005
|
| [4] |
Hamming I, Timens W, Bulthuis MLC, et al. (2004) Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J Pathol 203: 631-637. https://doi.org/10.1002/path.1570
|
| [5] | Williams S (2020) A brief history of human coronaviruses. The Scientist . Available from: https://www.the-scientist.com/news-opinion/a-brief-history-of-human-coronaviruses-67600. |
| [6] |
Fehr AR, Perlman S (2015) Coronaviruses: An overview of their replication and pathogenesis. Coronaviruses 1282: 1. https://doi.org/10.1007/978-1-4939-2438-7_1
|
| [7] |
Hulswit RJG, de Haan CAM, Bosch BJ (2016) Coronavirus spike protein and tropism changes. Adv Virus Res 96: 29-57. https://doi.org/10.1016/bs.aivir.2016.08.004
|
| [8] |
Chen L, Liu W, Zhang Q, et al. (2020) RNA based mNGS approach identifies a novel human coronavirus from two individual pneumonia cases in 2019 Wuhan outbreak. Emerg Microbes Infect 9: 313-319. https://doi.org/10.1080/22221751.2020.1725399
|
| [9] |
Hagemeijer MC, Verheije MH, Ulasli M, et al. (2010) Dynamics of coronavirus replication–transcription complexes. J Virol 84: 2134-2149. https://doi.org/10.1128/JVI.01716-09
|
| [10] |
Ruch TR, Machamer CE (2012) The coronavirus E protein: Assembly and beyond. Viruses 4: 363-382. https://doi.org/10.3390/v4030363
|
| [11] |
Beacon TH, Delcuve GP, Davie JR (2021) Epigenetic regulation of ACE2, the receptor of the SARS-CoV-2 virus. Genome 64: 386-399. https://doi.org/10.1139/gen-2020-0124
|
| [12] |
Neuman BW, Kiss G, Kunding AH, et al. (2011) A structural analysis of M protein in coronavirus assembly and morphology. J Struct Biol 174: 11-22. https://doi.org/10.1016/j.jsb.2010.11.021
|
| [13] |
Huang Y, Yang C, Xu X, et al. (2020) Structural and functional properties of SARS-CoV-2 spike protein: potential antivirus drug development for COVID-19. Acta Pharmacol Sin 41: 1141-1149. https://doi.org/10.1038/s41401-020-0485-4
|
| [14] |
V'kovski P, Kratzel A, Steiner S, et al. (2020) Coronavirus biology and replication: implications for SARS-CoV-2. Nat Rev Microbiol 19: 155-170. https://doi.org/10.1038/s41579-020-00468-6
|
| [15] |
Kumar M, Al Khodor S (2020) Pathophysiology and treatment strategies for COVID-19. J Transl Med 18: 353. https://doi.org/10.1186/s12967-020-02520-8
|
| [16] |
Chen Y, Liu Q, Guo D (2020) Emerging coronaviruses: Genome structure, replication, and pathogenesis. J Med Virol 92: 418-423. https://doi.org/10.1002/jmv.25681
|
| [17] | The Lancet Respiratory Medicine.COVID-19 transmission—up in the air. Lancet Respir Med (2020) 8: 1159. https://doi.org/10.1016/S2213-2600(20)30514-2 |
| [18] |
Cai J, Sun W, Huang J, et al. (2020) Indirect virus transmission in cluster of COVID-19 cases, Wenzhou, China, 2020. Emerg Infect Dis 26: 1343-1345. https://doi.org/10.3201/eid2606.200412
|
| [19] | CDC.Scientific brief: SARS-CoV-2 and potential airborne transmission. CDC (2021) . Available from: https://www.cdc.gov/coronavirus/2019-ncov/science/science-briefs/sars-cov-2-transmission.html. |
| [20] |
Lu R, Zhao X, Li J, et al. (2020) Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding. Lancet 395: 565-574. https://doi.org/10.1016/S0140-6736(20)30251-8
|
| [21] |
Schäfer A, Baric RS (2017) Epigenetic landscape during coronavirus infection. Pathogens 6: 8. https://doi.org/10.3390/pathogens6010008
|
| [22] |
Li F (2015) Receptor recognition mechanisms of coronaviruses: a decade of structural studies. J Virol 89: 1954-1964. https://doi.org/10.1128/JVI.02615-14
|
| [23] |
Atlante S, Mongelli A, Barbi V, et al. (2020) The epigenetic implication in coronavirus infection and therapy. Clin Epigenetics 12: 1-12. https://doi.org/10.1186/s13148-020-00946-x
|
| [24] |
Yan R, Zhang Y, Li Y, et al. (2020) Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 367: 1444-1448. https://doi.org/10.1126/science.abb2762
|
| [25] |
Tai W, He L, Zhang X, et al. (2020) Characterization of the receptor-binding domain (RBD) of 2019 novel coronavirus: implication for development of RBD protein as a viral attachment inhibitor and vaccine. Cell Mol Immunol 17: 613-620. https://doi.org/10.1038/s41423-020-0400-4
|
| [26] |
Clarke NE, Belyaev ND, Lambert DW, et al. (2014) Epigenetic regulation of angiotensin-converting enzyme 2 (ACE2) by SIRT1 under conditions of cell energy stress. Clin Sci 126: 507-516. https://doi.org/10.1042/CS20130291
|
| [27] | Muus C, Luecken MD, Eraslan G, et al. (2020) Integrated analyses of single-cell atlases reveal age, gender, and smoking status associations with cell type-specific expression of mediators of SARS-CoV-2 viral entry and highlights inflammatory programs in putative target cells. bioRxiv Preprint . |
| [28] |
Abdulhasan M, Ruden X, Rappolee B, et al. (2021) Stress decreases host viral resistance and increases covid susceptibility in embryonic stem cells. Stem Cell Rev Rep 17: 2164-2177. https://doi.org/10.1007/s12015-021-10188-w
|
| [29] |
Letko M, Marzi A, Munster V (2020) Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nat Microbiol 5: 562-569. https://doi.org/10.1038/s41564-020-0688-y
|
| [30] |
Lu R, Zhao X, Li J, et al. (2020) Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding. Lancet 395: 565-574. https://doi.org/10.1016/S0140-6736(20)30251-8
|
| [31] | da Silva SJR, de Lima SC, da Silva RC, et al. (2022) Viral load in COVID-19 patients: Implications for prognosis and vaccine efficacy in the context of emerging SARS-CoV-2 variants. Front Med 8: 3111. https://doi.org/10.3389/fmed.2021.836826 |
| [32] |
Paschos K, Allday MJ (2010) Epigenetic reprogramming of host genes in viral and microbial pathogenesis. Trends Microbiol 18: 439-447. https://doi.org/10.1016/j.tim.2010.07.003
|
| [33] |
Sawalha AH, Zhao M, Coit P, et al. (2020) Epigenetic dysregulation of ACE2 and interferon-regulated genes might suggest increased COVID-19 susceptibility and severity in lupus patients. Clin Immunol 215: 108410. https://doi.org/10.1016/j.clim.2020.108410
|
| [34] | Liebermanid NAP, Pedduid V, Xie H, et al. (2020) In vivo antiviral host transcriptional response to SARS-CoV-2 by viral load, sex, and age. PLoS Biol 18: e300084918. https://doi.org/10.1371/journal.pbio.3000849 |
| [35] |
Argyropoulos KV, Serrano A, Hu J, et al. (2020) Association of initial viral load in severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) patients with outcome and symptoms. Am J Pathol 190: 1881. https://doi.org/10.1016/j.ajpath.2020.07.001
|
| [36] |
Yuki K, Fujiogi M, Koutsogiannaki S (2020) COVID-19 pathophysiology: A review. Clin Immunol 215: 108427. https://doi.org/10.1016/j.clim.2020.108427
|
| [37] |
Cevik M, Kuppalli K, Kindrachuk J, et al. (2020) Virology, transmission, and pathogenesis of SARS-CoV-2. BMJ 371: m3862. https://doi.org/10.1136/bmj.m3862
|
| [38] |
Hojyo S, Uchida M, Tanaka K, et al. (2020) How COVID-19 induces cytokine storm with high mortality. Inflamm Regen 40: 37. https://doi.org/10.1186/s41232-020-00146-3
|
| [39] |
Obata Y, Furusawa Y, Hase K, et al. (2015) Epigenetic modifications of the immune system in health and disease. Immunol Cell Biol 93: 226-232. https://doi.org/10.1038/icb.2014.114
|
| [40] |
Jasiulionis MG (2018) Abnormal epigenetic regulation of immune system during aging. Front Immunol 9: 197. https://doi.org/10.3389/fimmu.2018.00197
|
| [41] |
Placek K, Schultze JL, Aschenbrenner AC (2019) Epigenetic reprogramming of immune cells in injury, repair, and resolution. J Clin Invest 129: 2994-3005. https://doi.org/10.1172/JCI124619
|
| [42] |
Cole J, Morris P, Dickman MJ, et al. (2016) The therapeutic potential of epigenetic manipulation during infectious diseases. Pharmacol Therapeut 167: 85-99. https://doi.org/10.1016/j.pharmthera.2016.07.013
|
| [43] |
Lynch KL, Gooding LR, Garnett-Benson C, et al. (2019) Epigenetics and the dynamics of chromatin during adenovirus infections. FEBS Lett 593: 3551-3570. https://doi.org/10.1002/1873-3468.13697
|
| [44] | Khan MAAK, Islam ABMMK (2020) SARS-CoV-2 proteins exploit host's genetic and epigenetic mediators for the annexation of key host signaling pathways that confers its immune evasion and disease pathophysiology. bioRxiv Preprint . https://doi.org/10.1101/2020.05.06.050260 |
| [45] |
Jantsch J, Schatz V, Friedrich D, et al. (2015) Cutaneous Na+ storage strengthens the antimicrobial barrier function of the skin and boosts macrophage-driven host defense. Cell Metab 21: 493-501. https://doi.org/10.1016/j.cmet.2015.02.003
|
| [46] |
Khansari N, Shakiba Y, Mahmoudi M (2009) Chronic inflammation and oxidative stress as a major cause of age- related diseases and cancer. Recent Pat Inflamm Allergy Drug Discov 3: 73-80. https://doi.org/10.2174/187221309787158371
|
| [47] |
Li X, Geng M, Peng Y, et al. (2020) Molecular immune pathogenesis and diagnosis of COVID-19. J Pharm Anal 10: 102-108. https://doi.org/10.1016/j.jpha.2020.03.001
|
| [48] |
Aung HT, Schroder K, Himes SR, et al. (2006) LPS regulates proinflammatory gene expression in macrophages by altering histone deacetylase expression. FASEB J 20: 1315-1327. https://doi.org/10.1096/fj.05-5360com
|
| [49] |
Villagra A, Cheng F, Wang HW, et al. (2009) The histone deacetylase HDAC11 regulates the expression of interleukin 10 and immune tolerance. Nat Immunol 10: 92-100. https://doi.org/10.1038/ni.1673
|
| [50] |
Jeudy S, Rigou S, Alempic JM, et al. (2020) The DNA methylation landscape of giant viruses. Nat Commun 11: 1-12. https://doi.org/10.1038/s41467-020-16414-2
|
| [51] |
Crimi E, Benincasa G, Figueroa-Marrero N, et al. (2020) Epigenetic susceptibility to severe respiratory viral infections and its therapeutic implications: a narrative review. Br J Anaesth 125: 1002-1017. https://doi.org/10.1016/j.bja.2020.06.060
|
| [52] |
Nicodeme E, Jeffrey KL, Schaefer U, et al. (2010) Suppression of inflammation by a synthetic histone mimic. Nature 468: 1119-1123. https://doi.org/10.1038/nature09589
|
| [53] |
Singh N, Gurav A, Sivaprakasam S, et al. (2014) Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity 40: 128-139. https://doi.org/10.1016/j.immuni.2013.12.007
|
| [54] |
Tsai K, Cullen BR (2020) Epigenetic and epitranscriptomic regulation of viral replication. Nat Rev Microbiol 18: 559-570. https://doi.org/10.1038/s41579-020-0382-3
|
| [55] |
Sinha P, Matthay MA, Calfee CS (2020) Is a ‘cytokine storm’ relevant to COVID-19?. JAMA Intern Med 180: 1152-1154. https://doi.org/10.1001/jamainternmed.2020.3313
|
| [56] |
Wiersinga WJ, Rhodes A, Cheng AC, et al. (2020) Pathophysiology, transmission, diagnosis, and treatment of coronavirus disease 2019 (COVID-19): A review. JAMA 324: 782-793. https://doi.org/10.1001/jama.2020.12839
|
| [57] |
Buchrieser J, Dufloo J, Hubert M, et al. (2020) Syncytia formation by SARS-CoV-2-infected cells. EMBO J 39: e106267. https://doi.org/10.15252/embj.2020106267
|
| [58] |
Ackermann M, Verleden SE, Kuehnel M, et al. (2020) Pulmonary vascular endothelialitis, thrombosis, and angiogenesis in COVID-19. N Engl J Med 383: 120-128. https://doi.org/10.1056/NEJMoa2015432
|
| [59] |
Jit BP, Qazi S, Arya R, et al. (2021) An immune epigenetic insight to COVID-19 infection. Epigenomics 13: 465-480. https://doi.org/10.2217/epi-2020-0349
|
| [60] |
Pervin Z, Hassan MM (2020) Synergistic therapeutic actions of antimicrobial peptides to treat multidrug-resistant bacterial infection. Rev Med Microbiol 32: 83-89. https://doi.org/10.1097/MRM.0000000000000239
|
| [61] |
Fischer N (2020) Infection-induced epigenetic changes and their impact on the pathogenesis of diseases. Semin Immunopathol 42: 127-130. https://doi.org/10.1007/s00281-020-00793-1
|
| [62] |
Paramasivam A, Priyadharsini JV (2020) Epigenetic modifications of RNA and their implications in antiviral immunity. Epigenomics 12: 1673-1675. https://doi.org/10.2217/epi-2020-0307
|
| [63] |
Singer BD, Mock JR, Aggarwal NR, et al. (2015) Regulatory T cell DNA methyltransferase inhibition accelerates resolution of lung inflammation. Am J Respir Cell Mol Biol 52: 641-652. https://doi.org/10.1165/rcmb.2014-0327OC
|
| [64] |
Fang J, Hao Q, Liu L, et al. (2012) Epigenetic changes mediated by microRNA miR29 activate cyclooxygenase 2 and lambda-1 interferon production during viral infection. J Virol 86: 1010-1020. https://doi.org/10.1128/JVI.06169-11
|
| [65] |
Huang KJ, Su IJ, Theron M, et al. (2005) An interferon-γ-related cytokine storm in SARS patients. J Med Virol 75: 185-194. https://doi.org/10.1002/jmv.20255
|
| [66] |
Stergioti EM, Manolakou T, Boumpas DT, et al. (2022) Antiviral innate immune responses in autoimmunity: Receptors, pathways, and therapeutic targeting. Biomedicines 10: 2820. https://doi.org/10.3390/biomedicines10112820
|
| [67] |
Wu C, Chen X, Cai Y, et al. (2020) Risk factors associated with acute respiratory distress syndrome and death in patients with coronavirus disease 2019 pneumonia in Wuhan, China. JAMA Intern Med 180: 934-943. https://doi.org/10.1001/jamainternmed.2020.0994
|
| [68] |
Ellinghaus D, Degenhardt F, Bujanda L, et al. (2020) Genomewide association study of severe COVID-19 with respiratory failure. N Engl J Med 383: 1522-1534. https://doi.org/10.1056/NEJMoa2020283
|
| [69] | Ellinghaus D, Degenhardt F, Bujanda L, et al. (2020) The ABO blood group locus and a chromosome 3 gene cluster associate with SARS-CoV-2 respiratory failure in an Italian-Spanish genome-wide association analysis. medRxiv Preprint . |
| [70] |
Pairo-Castineira E, Clohisey S, Klaric L, et al. (2021) Genetic mechanisms of critical illness in COVID-19. Nature 591: 92-98. https://doi.org/10.1038/s41586-020-03065-y
|
| [71] |
Zeberg H, Pääbo S (2020) The major genetic risk factor for severe COVID-19 is inherited from Neanderthals. Nature 587: 610-612. https://doi.org/10.1038/s41586-020-2818-3
|
| [72] |
Mueller AL, Mcnamara MS, Sinclair DA (2020) Why does COVID-19 disproportionately affect older people?. Aging 12: 9959-9981. https://doi.org/10.18632/aging.103344
|
| [73] |
Senapati S, Kumar S, Singh AK, et al. (2020) Assessment of risk conferred by coding and regulatory variations of TMPRSS2 and CD26 in susceptibility to SARS-CoV-2 infection in human. J Genet 99: 1-5. https://doi.org/10.1007/s12041-020-01217-7
|
| [74] |
Wimalawansa SJ (2019) Vitamin D deficiency: Effects on oxidative stress, epigenetics, gene regulation, and aging. Biology 8: 30. https://doi.org/10.3390/biology8020030
|
| [75] |
Chen L, Dong Y, Bhagatwala J, et al. (2019) Effects of vitamin D3 supplementation on epigenetic aging in overweight and obese African Americans with suboptimal vitamin D status: A randomized clinical trial. J Gerontol A-Biol 74: 91. https://doi.org/10.1093/gerona/gly223
|
| [76] |
Zhang Y, Zhang Y, Qin L, et al. (2020) Interferon-induced transmembrane protein 3 genetic variant rs12252-C associated with disease severity in coronavirus disease 2019. J Infect Dis 222: 34-37. https://doi.org/10.1093/infdis/jiaa224
|
| [77] |
Li S, Ma F, Yokota T, et al. (2021) Metabolic reprogramming and epigenetic changes of vital organs in SARS-CoV-2–induced systemic toxicity. JCI Insight 6: e145027. https://doi.org/10.1172/jci.insight.145027
|
| [78] |
Gupta A, Madhavan MV, Sehgal K, et al. (2020) Extrapulmonary manifestations of COVID-19. Nat Med 26: 1017-1032. https://doi.org/10.1038/s41591-020-0968-3
|
| [79] |
Channappanavar R, Fehr AR, Vijay R, et al. (2016) Dysregulated type I interferon and inflammatory monocyte-macrophage responses cause lethal pneumonia in SARS-CoV-infected mice. Cell Host Microbe 19: 181-193. https://doi.org/10.1016/j.chom.2016.01.007
|
| [80] |
Anderson G, Maes M (2020) Gut dysbiosis dysregulates central and systemic homeostasis via suboptimal mitochondrial function: Assessment, treatment and classification implications. Curr Top Med Chem 20: 524-539. https://doi.org/10.2174/1568026620666200131094445
|
| [81] |
Mika A, Rumian N, Loughridge AB, et al. (2016) Exercise and prebiotics produce stress resistance: Converging impacts on stress-protective and butyrate-producing gut bacteria. Int Rev Neurobiol 131: 165-191. https://doi.org/10.1016/bs.irn.2016.08.004
|
| [82] |
Anderson G, Reiter RJ (2020) Melatonin: Roles in influenza, COVID-19, and other viral infections. Rev Med Virol 30: e2109. https://doi.org/10.1002/rmv.2109
|
| [83] |
Jin CJ, Engstler AJ, Sellmann C, et al. (2016) Sodium butyrate protects mice from the development of the early signs of non-alcoholic fatty liver disease: role of melatonin and lipid peroxidation. Br J Nutr 116: 1682-1693. https://doi.org/10.1017/S0007114516004025
|
| [84] |
Channappanavar R, Perlman S (2017) Pathogenic human coronavirus infections: causes and consequences of cytokine storm and immunopathology. Semin Immunopathol 39: 529-539. https://doi.org/10.1007/s00281-017-0629-x
|
| [85] |
Acosta PL, Byrne AB, Hijano DR, et al. (2020) Human type I interferon antiviral effects in respiratory and reemerging viral infections. J Immunol Res 2020: 1372494. https://doi.org/10.1155/2020/1372494
|
| [86] |
Barrett S, Goh J, Coughlan B, et al. (2001) The natural course of hepatitis C virus infection after 22 years in a unique homogenous cohort: spontaneous viral clearance and chronic HCV infection. Gut 49: 423-430. https://doi.org/10.1136/gut.49.3.423
|
| [87] |
Bogdanović Z, Marinović-Terzić I, Kuret S, et al. (2016) The impact of IL-6 and IL-28B gene polymorphisms on treatment outcome of chronic hepatitis C infection among intravenous drug users in Croatia. PeerJ 2016: e2576. https://doi.org/10.7717/peerj.2576
|
| [88] |
Nattermann J, Vogel M, Nischalke HD, et al. (2008) The transforming growth factor-beta high-producer genotype is associated with response to hepatitis C virus-specific therapy in HIV-positive patients with acute hepatitis C. AIDS 22: 1287-1292. https://doi.org/10.1097/QAD.0b013e3282f85daa
|
| [89] |
Beltrán-García J, Osca-Verdegal R, Pallardó FV, et al. (2020) Oxidative stress and inflammation in COVID-19-associated sepsis: The potential role of anti-oxidant therapy in avoiding disease progression. Antioxidants 9: 1-20. https://doi.org/10.3390/antiox9100936
|
| [90] | Wang F, Huang S, Gao R, et al. (2020) Initial whole-genome sequencing and analysis of the host genetic contribution to COVID-19 severity and susceptibility. Cell Discov 6: 1-16. https://doi.org/10.1038/s41421-020-00231-4 |
| [91] | Cao X, Li W, Wang T, et al. (2022) Accelerated biological aging in COVID-19 patients. Nat Commun 13: 1-7. https://doi.org/10.1038/s41467-022-29801-8 |
| [92] |
Li X, Zhong X, Wang Y, et al. (2021) Clinical determinants of the severity of COVID-19: A systematic review and meta-analysis. PLoS One 16: e0250602. https://doi.org/10.1371/journal.pone.0250602
|
| [93] |
Fu J, Reid SA, French B, et al. (2022) Racial disparities in COVID-19 outcomes among black and white patients with cancer. JAMA Netw Open 5: e224304. https://doi.org/10.1001/jamanetworkopen.2022.4304
|
| [94] | Gu T, Mack JA, Salvatore M, et al. (2020) COVID-19 outcomes, risk factors and associations by race: a comprehensive analysis using electronic health records data in Michigan Medicine. medRxiv Preprint . https://doi.org/10.1101/2020.06.16.20133140 |
| [95] | Aman F, Masood S (2020) How nutrition can help to fight against COVID-19 pandemic. Pakistan J Med Sci 36: S121. https://doi.org/10.12669/pjms.36.COVID19-S4.2776 |
| [96] |
Merino J, Joshi AD, Nguyen LH, et al. (2021) Diet quality and risk and severity of COVID-19: a prospective cohort study. Gut 70: 2096-2104. https://doi.org/10.1136/gutjnl-2021-325353
|
| [97] | Kuehn BM (2021) More severe obesity leads to more severe COVID-19 in study. JAMA 325: 1603-1603. https://doi.org/10.1001/jama.2021.4853 |
| [98] | Beaumont Health.How obesity impacts your risk for severe COVID-19. Beaumont Health (2022) . Available from: https://www.beaumont.org/health-wellness/blogs/how-obesity-impacts-your-risk-for-severe-covid-19. |
| [99] |
Wieczfinska J, Kleniewska P, Pawliczak R (2022) Oxidative stress-related mechanisms in SARS-CoV-2 infections. Oxid Med Cell Longev 2022: 5589089. https://doi.org/10.1155/2022/5589089
|
| [100] |
Yek C, Warner S, Mancera A, et al. (2022) Misclassification bias in estimating clinical severity of SARS-CoV-2 variants. Lancet 400: 809. https://doi.org/10.1016/S0140-6736(22)01469-6
|
| [101] |
Liu BM, Hill HR (2020) Role of host immune and inflammatory responses in COVID-19 cases with underlying primary immunodeficiency: A review. J Interferon Cytokine Res 40: 549-554. https://doi.org/10.1089/jir.2020.0210
|
| [102] |
St Sauver JL, Lopes GS, Rocca WA, et al. (2021) Factors associated with severe COVID-19 infection among persons of different ages living in a defined midwestern US population. Mayo Clin Proc 96: 2528-2539. https://doi.org/10.1016/j.mayocp.2021.06.023
|
| [103] |
Pearce EL, Pearce EJ (2013) Metabolic pathways in immune cell activation and quiescence. Immunity 38: 633. https://doi.org/10.1016/j.immuni.2013.04.005
|
| [104] |
Wang QQ, Kaelber DC, Xu R, et al. (2021) COVID-19 risk and outcomes in patients with substance use disorders: analyses from electronic health records in the United States. Mol Psychiatr 26: 30-39. https://doi.org/10.1038/s41380-020-00880-7
|
| [105] |
Clift AK, von Ende A, Tan PS, et al. (2022) Smoking and COVID-19 outcomes: an observational and Mendelian randomisation study using the UK Biobank cohort. Thorax 77: 65-73. https://doi.org/10.1136/thoraxjnl-2021-217080
|
| [106] |
Polverino F (2020) Cigarette smoking and COVID-19: A complex interaction. Am J Resp Crit Care 202: 471-472. https://doi.org/10.1164/rccm.202005-1646LE
|
| [107] |
Pervin Z, Stephen JM (2021) Effect of alcohol on the central nervous system to develop neurological disorder: pathophysiological and lifestyle modulation can be potential therapeutic options for alcohol-induced neurotoxication. AIMS Neurosci 8: 390-413. https://doi.org/10.3934/Neuroscience.2021021
|
| [108] | Rodríguez CV, Horcajadas FA, Arroba CMA, et al. (2021) COVID-19-related neuropsychiatric symptoms in patients with alcohol abuse conditions during the SARS-CoV-2 pandemic: A retrospective cohort study using real world data from electronic health records of a tertiary hospital. Front Neurol 12: 162. https://doi.org/10.3389/fneur.2021.630566 |
| [109] |
Netea MG, Giamarellos-Bourboulis EJ, Domínguez-Andrés J, et al. (2020) Trained immunity: a tool for reducing susceptibility to and the severity of SARS-CoV-2 infection. Cell 181: 969-977. https://doi.org/10.1016/j.cell.2020.04.042
|
| [110] |
Kerboua KE (2020) The perplexing question of trained immunity vs adaptive memory in COVID-19. J Med Virol 92: 1858-1863. https://doi.org/10.1002/jmv.26083
|
| [111] | Rabaan AA, Al Mutair A, Alhumaid S, et al. (2021) Modulation of host epigenome by coronavirus infections and developing treatment modalities for COVID-19 beyond genetics. Eur Rev Med Pharmacol Sci 25: 5947-5964. |
| [112] |
Su Q, Wang S, Baltzis D, et al. (2007) Interferons induce tyrosine phosphorylation of the eIF2α kinase PKR through activation of Jak1 and Tyk2. EMBO Rep 8: 265. https://doi.org/10.1038/sj.embor.7400891
|
| [113] |
El Baba R, Herbein G (2020) Management of epigenomic networks entailed in coronavirus infections and COVID-19. Clin Epigenetics 12: 118. https://doi.org/10.1186/s13148-020-00912-7
|

Figures(2) / Tables(1)

Zinia Pervin, Anika Tasnim, Hasib Ahamed, Md Al Hasibuzzaman. Epigenetic regulation of the COVID-19 pathogenesis: its impact on the host immune response and disease progression[J]. AIMS Allergy and Immunology, 2023, 7(1): 60-81. doi: 10.3934/Allergy.2023005







 DownLoad:
DownLoad: