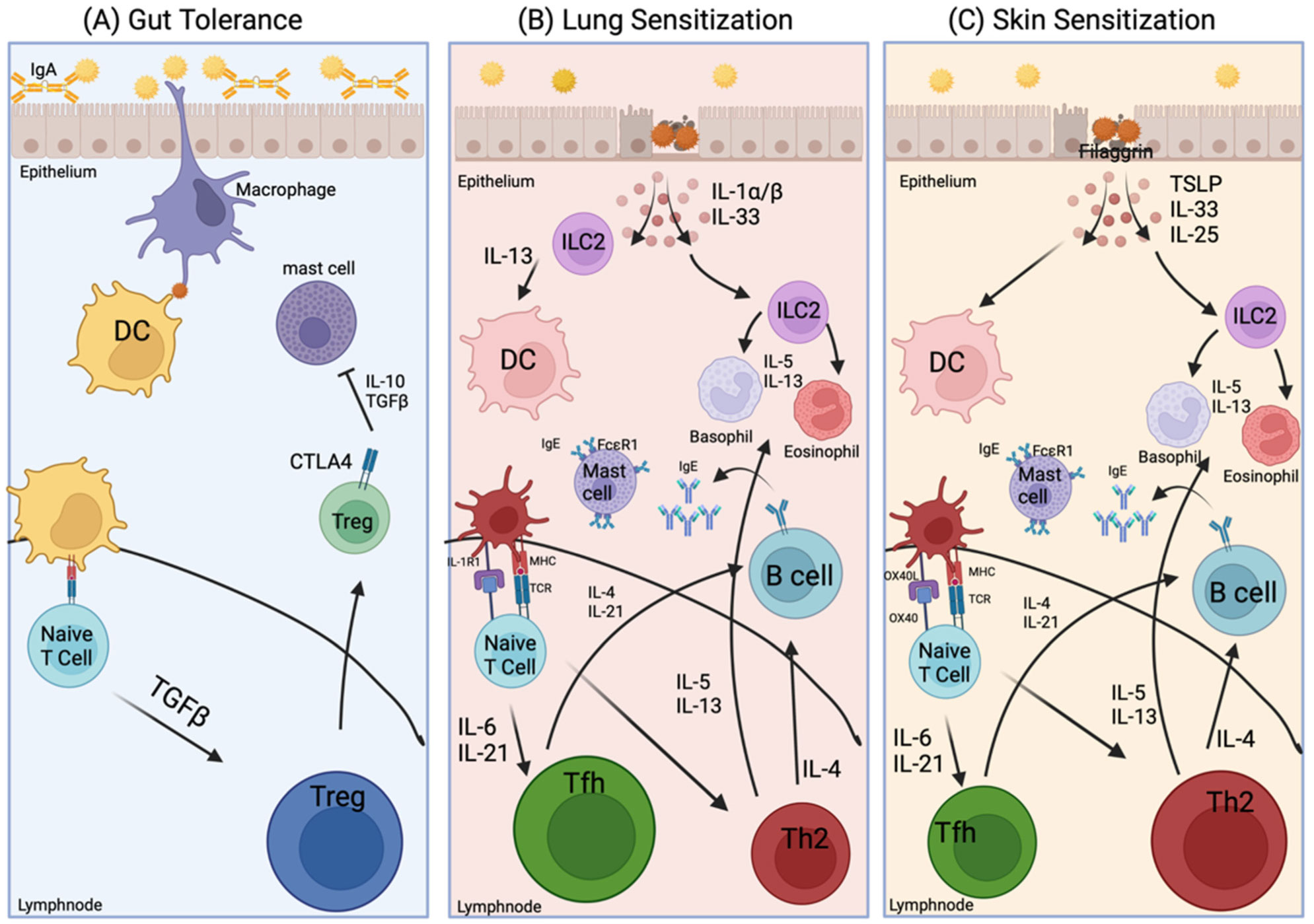
Food allergies are of great public health concern due to their rising prevalence. Our understanding of how the immune system reacts to food remains incomplete. Allergic responses vary between individuals with food allergies. This variability could be caused by genetic, environmental, hormonal, or metabolic factors that impact immune responses mounted against allergens found in foods. Peanut (PN) allergy is one of the most severe and persistent of food allergies, warranting examination into how sensitization occurs to drive IgE-mediated allergic reactions. In recent years, much has been learned about the mechanisms behind the initiation of IgE-mediated food allergies, but additional questions remain. One unresolved issue is whether sex hormones impact the development of food allergies. Sex differences are known to exist in other allergic diseases, so this poses the question about whether the same phenomenon is occurring in food allergies. Studies show that females exhibit a higher prevalence of atopic conditions, such as allergic asthma and eczema, relative to males. Discovering such sex differences in allergic diseases provide a basis for investigating the mechanisms of how hormones influence the development of IgE-mediated reactions to foods. Analysis of existing food allergy demographics found that they occur more frequently in male children and adult females, which is comparable to allergic asthma. This paper reviews existing allergic mechanisms, sensitization routes, as well as how sex hormones may play a role in how the immune system reacts to common food allergens such as PN.
Citation: McKenna S. Vininski, Sunanda Rajput, Nicholas J. Hobbs, Joseph J. Dolence. Understanding sex differences in the allergic immune response to food[J]. AIMS Allergy and Immunology, 2022, 6(3): 90-105. doi: 10.3934/Allergy.2022009
Food allergies are of great public health concern due to their rising prevalence. Our understanding of how the immune system reacts to food remains incomplete. Allergic responses vary between individuals with food allergies. This variability could be caused by genetic, environmental, hormonal, or metabolic factors that impact immune responses mounted against allergens found in foods. Peanut (PN) allergy is one of the most severe and persistent of food allergies, warranting examination into how sensitization occurs to drive IgE-mediated allergic reactions. In recent years, much has been learned about the mechanisms behind the initiation of IgE-mediated food allergies, but additional questions remain. One unresolved issue is whether sex hormones impact the development of food allergies. Sex differences are known to exist in other allergic diseases, so this poses the question about whether the same phenomenon is occurring in food allergies. Studies show that females exhibit a higher prevalence of atopic conditions, such as allergic asthma and eczema, relative to males. Discovering such sex differences in allergic diseases provide a basis for investigating the mechanisms of how hormones influence the development of IgE-mediated reactions to foods. Analysis of existing food allergy demographics found that they occur more frequently in male children and adult females, which is comparable to allergic asthma. This paper reviews existing allergic mechanisms, sensitization routes, as well as how sex hormones may play a role in how the immune system reacts to common food allergens such as PN.

| [1] |
Jiang J, Bushara O, Ponczek J, et al. (2018) Updated pediatric peanut allergy prevalence in the United States. Ann Allerg Asthma Im 121: S14. https://doi.org/10.1016/j.anai.2018.09.042 
|
| [2] |
Gupta RS, Warren CM, Smith BM, et al. (2019) Prevalence and severity of food allergies among US adults. JAMA Netw Open 2: e185630-e185630. https://doi.org/10.1001/jamanetworkopen.2018.5630
|
| [3] |
Lieberman JA, Gupta RS, Knibb RC, et al. (2021) The global burden of illness of peanut allergy: A comprehensive literature review. Allergy 76: 1367-1384. https://doi.org/10.1111/all.14666
|
| [4] |
Togias A, Cooper SF, Acebal ML, et al. (2017) Addendum guidelines for the prevention of peanut allergy in the United States: Report of the National Institute of Allergy and Infectious Diseases—sponsored expert panel. J Allergy Clin Immun 139: 29-44. https://doi.org/10.1016/j.jaci.2016.10.010
|
| [5] |
Dolence JJ, Kita H (2020) Allergic sensitization to peanuts is enhanced in mice fed a high-fat diet. AIMS Allerg Immu 4: 88-99. https://doi.org/10.3934/Allergy.2020008
|
| [6] |
Zhu TH, Zhu TR, Tran KA, et al. (2018) Epithelial barrier dysfunctions in atopic dermatitis: a skin–gut–lung model linking microbiome alteration and immune dysregulation. Brit J Dermatol 179: 570-581. https://doi.org/10.1111/bjd.16734
|
| [7] |
Zhu Y, Shao X, Wang X, et al. (2019) Sex disparities in cancer. Cancer Lett 466: 35-38. https://doi.org/10.1016/j.canlet.2019.08.017
|
| [8] |
Fuseini H, Newcomb DC (2017) Mechanisms driving gender differences in asthma. Physiol Behav 176: 139-148. https://doi.org/10.1007/s11882-017-0686-1
|
| [9] |
Gubbels Bupp MR, Jorgensen TN (2018) Androgen-induced immunosuppression. Front Immunol 9: 794. https://doi.org/10.3389/fimmu.2018.00794
|
| [10] |
Warren C, Lei D, Sicherer S, et al. (2021) Prevalence and characteristics of peanut allergy in US adults. J Allergy Clin Immun 147: 2263-2270. https://doi.org/10.1016/j.jaci.2020.11.046
|
| [11] | Hernández-Colín DD, Fregoso-Zúñiga E, Morales-Romero J, et al. (2019) Peanut allergy among Mexican adults with allergic respiratory diseases: prevalence and clinical manifestations. Rev Alerg Mex 66: 314-321. https://doi.org/10.29262/ram.v66i3.619 |
| [12] |
Dolence JJ, Kobayashi T, Iijima K, et al. (2018) Airway exposure initiates peanut allergy by involving the IL-1 pathway and T follicular helper cells in mice. J Allergy Clin Immun 142: 1144-1158. https://doi.org/10.1016/j.jaci.2017.11.020
|
| [13] |
Du Toit G, Roberts G, Sayre PH, et al. (2015) Randomized trial of peanut consumption in infants at risk for peanut allergy. N Engl J Med 372: 803-813. https://doi.org/10.1056/NEJMoa1414850
|
| [14] |
Du Toit G, Katz Y, Sasieni P, et al. (2008) Early consumption of peanuts in infancy is associated with a low prevalence of peanut allergy. J Allergy Clin Immun 122: 984-991. https://doi.org/10.1016/j.jaci.2008.08.039
|
| [15] |
Platts-Mills TAE (2001) The role of immunoglobulin E in allergy and asthma. Am J Resp Crit Care 164: S1-S5. https://doi.org/10.1164/ajrccm.164.supplement_1.2103024
|
| [16] | Fuller RW, Morris PK, Richmond R, et al. (1986) Immunoglobulin E-dependent stimulation of human alveolar macrophages: Significant in type 1 hypersensitivity. Clin Exp Immunol 65: 416-426. |
| [17] |
Viel S, Pescarmona R, Belot A, et al. (2018) A case of type 2 hypersensitivity to rasburicase diagnosed with a Natural Killer cell activation assay. Front Immunol 9: 110. https://doi.org/10.3389/fimmu.2018.00110
|
| [18] | Eggleton P Hypersensitivity: Immune Complex Mediated (Type III) (2013). eLS |
| [19] |
Uzzaman A, Cho SH (2012) Classification of hypersensitivity reactions. Allergy Asthma Proc 33: 96-99. https://doi.org/10.2500/aap.2012.33.3561
|
| [20] |
Sampson HA, O'Mahony L, Burks AW, et al. (2018) Mechanisms of food allergy. J Allergy Clin Immun 141: 11-19. https://doi.org/10.1016/j.jaci.2017.11.005
|
| [21] |
Pali-Schöll I, Herzog R, Wallmann J, et al. (2010) Antacids and dietary supplements with an influence on the gastric pH increase the risk for food sensitization. Clin Exp Allergy 40: 1091-1098. https://doi.org/10.1111/j.1365-2222.2010.03468.x
|
| [22] |
Van Erp FC, Knulst AC, Meijer Y, et al. (2014) Standardized food challenges are subject to variability in interpretation of clinical symptoms. Clin Transl Allergy 4: 1-6. https://doi.org/10.1186/s13601-014-0043-6
|
| [23] |
Dyer AA, Rivkina V, Perumal D, et al. (2015) Epidemiology of childhood peanut allergy. Allergy Asthma Proc 36: 58-64. https://doi.org/10.2500/aap.2015.36.3819
|
| [24] | Gupta RS, Warren C, Smith BM, et al. (2019) The public health impact of parent-reported childhood food allergies in the United States. Pediatrics 144: S28. https://doi.org/10.1542/peds.2019-2461PP |
| [25] |
Sicherer SH, Morrow EH, Sampson HA (2000) Dose-response in double-blind, placebo-controlled oral food challenges in children with atopic dermatitis. J Allergy Clin Immun 105: 582-586. https://doi.org/10.1067/mai.2000.104941
|
| [26] |
Du Toit G, Roberts G, Sayre PH, et al. (2015) Randomized trial of peanut consumption in infants at risk for peanut allergy. N Engl J Med 372: 803-813. https://doi.org/10.1056/NEJMoa1414850
|
| [27] |
Smeekens JM, Immormino RM, Balogh PA, et al. (2019) Indoor dust acts as an adjuvant to promote sensitization to peanut through the airway. Clin Exp Allergy 49: 1500-1511. https://doi.org/10.1111/cea.13486
|
| [28] |
Trendelenburg V, Ahrens B, Wehrmann AK, et al. (2013) Peanut allergen in house dust of eating area and bed—A risk factor for peanut sensitization?. Allergy 68: 1460-1462. https://doi.org/10.1111/all.12226
|
| [29] |
Anvari S, Miller J, Yeh CY, et al. (2019) IgE-mediated food allergy. Clin Rev Allerg Immu 57: 244-260. https://doi.org/10.1007/s12016-018-8710-3
|
| [30] |
Gri G, Piconese S, Frossi B, et al. (2008) CD4+CD25+ regulatory T cells suppress mast cell degranulation and allergic responses through OX40-OX40L interaction. Immunity 29: 771-781. https://doi.org/10.1016/j.immuni.2008.08.018
|
| [31] |
Tordesillas L, Berin MC (2018) Mechanisms of oral tolerance. Clin Rev Allerg Immu 55: 107-117. https://doi.org/10.1007/s12016-018-8680-5
|
| [32] |
Goleva E, Berdyshev E, Leung DYM (2019) Epithelial barrier repair and prevention of allergy. J Clin Invest 129: 1463-1474. https://doi.org/10.1172/JCI124608
|
| [33] |
Eiwegger T, Hung L, San Diego KE, et al. (2019) Recent developments and highlights in food allergy. Allergy 74: 2355-2367. https://doi.org/10.1111/all.14082
|
| [34] |
Platts-Mills TAE, Woodfolk JA (2011) Allergens and their role in the allergic immune response. Immunol Rev 242: 51-68. https://doi.org/10.1111/j.1600-065X.2011.01021.x
|
| [35] |
Heib V, Becker M, Taube C, et al. (2008) Advances in the understanding of mast cell function. Brit J Haematol 142: 683-694. https://doi.org/10.1111/j.1365-2141.2008.07244.x
|
| [36] |
Krempski JW, Kobayashi T, Iijima K, et al. (2020) Group 2 innate lymphoid cells promote development of T follicular helper cells and initiate allergic sensitization to peanuts. J Immunol 204: 3086-3096. https://doi.org/10.4049/jimmunol.2000029
|
| [37] |
Tordesillas L, Goswami R, Benedé S, et al. (2014) Skin exposure promotes a Th2-dependent sensitization to peanut allergens. J Clin Invest 124: 4965-4975. https://doi.org/10.1172/JCI75660
|
| [38] | Krempski JW, Lama JK, Iijima K, et al. (2022) A mouse model of the “LEAP” study reveals a role for CTLA-4 in preventing peanut allergy induced by environmental peanut exposure. J Allergy Clin Immun . In press. https://doi.org/10.1016/j.jaci.2022.02.024 |
| [39] |
Kulis MD, Smeekens JM, Immormino RM, et al. (2021) The airway as a route of sensitization to peanut: An update to the dual allergen exposure hypothesis. J Allergy Clin Immun 148: 689-693. https://doi.org/10.1016/j.jaci.2021.05.035
|
| [40] |
Brough HA, Santos AF, Makinson K, et al. (2013) Peanut protein in household dust is related to household peanut consumption and is biologically active. J Allergy Clin Immun 132: 630-638. https://doi.org/10.1016/j.jaci.2013.02.034
|
| [41] |
Flohr C, England K, Radulovic S, et al. (2010) Filaggrin loss-of-function mutations are associated with early-onset eczema, eczema severity and transepidermal water loss at 3 months of age. Brit J Dermatol 163: 1333-1336. https://doi.org/10.1111/j.1365-2133.2010.10068.x
|
| [42] |
Brough HA, Simpson A, Makinson K, et al. (2014) Peanut allergy: Effect of environmental peanut exposure in children with filaggrin loss-of-function mutations. J Allergy Clin Immun 134: 867-875. https://doi.org/10.1016/j.jaci.2014.08.011
|
| [43] |
Venkataraman D, Soto-Ramírez N, Kurukulaaratchy RJ, et al. (2014) Filaggrin loss-of-function mutations are associated with food allergy in childhood and adolescence. J Allergy Clin Immun 134: 876-882. https://doi.org/10.1016/j.jaci.2014.07.033
|
| [44] |
Brown SJ, Asai Y, Cordell HJ, et al. (2011) Loss-of-function variants in the filaggrin gene are a significant risk factor for peanut allergy. J Allergy Clin Immun 127: 661-667. https://doi.org/10.1016/j.jaci.2011.01.031
|
| [45] |
Wang YH (2016) Developing food allergy: A potential immunologic pathway linking skin barrier to gut. F1000Research 5: 1-8. https://doi.org/10.12688/f1000research.9497.1
|
| [46] |
Tordesillas L, Berin MC, Sampson HA (2017) Immunology of food allergy. Immunity 47: 32-50. https://doi.org/10.1016/j.immuni.2017.07.004
|
| [47] |
Ito T, Wang YH, Duramad O, et al. (2005) TSLP-activated dendritic cells induce an inflammatory T helper type 2 cell response through OX40 ligand. J Exp Med 202: 1213-1223. https://doi.org/10.1084/jem.20051135
|
| [48] |
Staden U, Rolinck-Werninghaus C, Brewe F, et al. (2007) Specific oral tolerance induction in food allergy in children: Efficacy and clinical patterns of reaction. Allergy 62: 1261-1269. https://doi.org/10.1111/j.1398-9995.2007.01501.x
|
| [49] |
Logan K, Du Toit G, Giovannini M, et al. (2020) Pediatric allergic diseases, food allergy, and oral tolerance. Annu Rev Cell Dev Bi 36: 511-528. https://doi.org/10.1146/annurev-cellbio-100818-125346
|
| [50] |
Mccallister JW, Mastronarde JG (2008) Sex differences in asthma. J Asthma 45: 853-861. https://doi.org/10.1080/02770900802444187
|
| [51] |
Schatz M, Clark S, Camargo CA (2006) Sex differences in the presentation and course of asthma hospitalizations. Chest 129: 50-55. https://doi.org/10.1378/chest.129.1.50
|
| [52] |
Klein SL, Flanagan KL (2016) Sex differences in immune responses. Nat Rev Immunol 16: 626-638. https://doi.org/10.1038/nri.2016.90
|
| [53] |
Furman D, Hejblum BP, Simon N, et al. (2014) Systems analysis of sex differences reveals an immunosuppressive role for testosterone in the response to influenza vaccination. P Natl Acad Sci USA 111: 869-874. https://doi.org/10.1073/pnas.1321060111
|
| [54] |
Keselman A, Heller N (2015) Estrogen signaling modulates allergic inflammation and contributes to sex differences in asthma. Front Immunol 6: 568. https://doi.org/10.3389/fimmu.2015.00568
|
| [55] |
Gilliver SC (2010) Sex steroids as inflammatory regulators. J Steroid Biochem 120: 105-115. https://doi.org/10.1016/j.jsbmb.2009.12.015
|
| [56] |
Afify SM, Pali-Schöll I (2017) Adverse reactions to food: The female dominance—A secondary publication and update. World Allergy Organ J 10: 1-8. https://doi.org/10.1186/s40413-017-0174-z
|
| [57] |
Fink AL, Klein SL (2018) The evolution of greater humoral immunity in females than males: implications for vaccine efficacy. Curr Opin Physiol 6: 16-20. https://doi.org/10.1016/j.cophys.2018.03.010
|
| [58] |
Becklake MR, Kauffmann F (1999) Gender differences in airway behaviour over the human life span. Thorax 54: 1119-1138. https://doi.org/10.1136/thx.54.12.1119
|
| [59] |
Fu L, Freishtat RJ, Gordish-Dressman H, et al. (2014) Natural progression of childhood asthma symptoms and strong influence of sex and puberty. Ann Am Thorac Soc 11: 898-907. https://doi.org/10.1513/AnnalsATS.201402-084OC
|
| [60] |
Carey MA, Card JW, Voltz JW, et al. (2007) It's all about sex: gender, lung development and lung disease. Trends Endocrin Met 18: 308-313. https://doi.org/10.1016/j.tem.2007.08.003
|
| [61] |
DunnGalvin A, Hourihane JOB, Frewer L, et al. (2006) Incorporating a gender dimension in food allergy research: A review. Allergy 61: 1336-1343. https://doi.org/10.1111/j.1398-9995.2006.01181.x
|
| [62] |
Agarwal AK, Shah A (1997) Menstrual-linked asthma. J Asthma 34: 539-545. https://doi.org/10.3109/02770909709055398
|
| [63] |
Murphy VE, Clifton VL, Gibson PG (2006) Asthma exacerbations during pregnancy: Incidence and association with adverse pregnancy outcomes. Thorax 61: 169-176. https://doi.org/10.1136/thx.2005.049718
|
| [64] |
Rao CK, Moore CG, Bleecker E, et al. (2013) Characteristics of perimenstrual asthma and its relation to asthma severity and control: Data from the Severe Asthma Research Program. Chest 143: 984-992. https://doi.org/10.1378/chest.12-0973
|
| [65] |
Schatz M, Harden K, Forsythe A, et al. (1988) The course of asthma during pregnancy, post partum, and with successive pregnancies: A prospective analysis. J Allergy Clin Immun 81: 495-504. https://doi.org/10.1016/0091-6749(88)90187-X
|
| [66] |
Shames RS, Heilbron DC, Janson SL, et al. (1998) Clinical differences among women with and without self-reported perimenstrual asthma. Ann Allerg Asthma Im 81: 65-72. https://doi.org/10.1016/S1081-1206(10)63111-0
|
| [67] |
Stenius-Aarniala B, Piirila P, Teramo K (1988) Asthma and pregnancy: a prospective study of 198 pregnancies. Thorax 43: 12-18. https://doi.org/10.1136/thx.43.1.12
|
| [68] |
Pali-Schöll I, Jensen-Jarolim E (2019) Gender aspects in food allergy. Curr Opin Allergy Clin Immunol 19: 249-255. https://doi.org/10.1097/ACI.0000000000000529
|
| [69] |
Bender BYAE, Matthews R (1981) Adverse foods. Brit J Nutr 46: 403-407. https://doi.org/10.1079/BJN19810048
|
| [70] |
Burr ML, Merrett TG (1983) Food intolerance: a community survey. Brit J Nutr 49: 217-219. https://doi.org/10.1079/BJN19830028
|
| [71] | Metcalfe D, Sampson H, Simon RA, et al. (2008) Food Allergy: Adverse Reactions to Foods and Food Additives. Malden: Blackwell Pub. https://doi.org/10.1002/9781444300062 |
| [72] |
Moneret-Vautrin DA, Morisset M (2005) Adult food allergy. Curr Allergy Asthma Rep 5: 80-85. https://doi.org/10.1007/s11882-005-0060-6
|
| [73] |
Schäfer T, Böhler E, Ruhdorfer S, et al. (2001) Epidemiology of food allergy/food intolerance in adults: associations with other manifestations of atopy. Allergy 56: 1172-1179. https://doi.org/10.1034/j.1398-9995.2001.00196.x
|
| [74] |
Jansen JJ, Kardinaal AF, Huijbers G, et al. (1994) Prevalence of food allergy and intolerance in the adult Dutch population. J Allergy Clin Immun 93: 446-456. https://doi.org/10.1016/0091-6749(94)90353-0
|
| [75] |
Young E, Stoneham MD, Petruckevitch A, et al. (1994) A population study of food intolerance. Lancet 343: 1127-1130. https://doi.org/10.1016/S0140-6736(94)90234-8
|
| [76] | Løvik M, Namork E, Fæste C, et al. (2009) The Norwegian national reporting system and register of severe allergic reactions to food. Nor Epidemiol 14: 155-160. https://doi.org/10.5324/nje.v14i2.238 |
| [77] |
Kelly C, Gangur V (2009) Sex disparity in food allergy: evidence from the PubMed Database. J Allergy 2009: 1-7. https://doi.org/10.1155/2009/159845
|
| [78] |
Mohammad I, Starskaia I, Nagy T, et al. (2018) Estrogen receptor contributes to T cell-mediated autoimmune inflammation by promoting T cell activation and proliferation. Sci Signal 11: 1-13. https://doi.org/10.1126/scisignal.aap9415
|
| [79] |
Heitkemper MM, Chang L (2009) Do fluctuations in ovarian hormones affect gastrointestinal symptoms in women with irritable bowel syndrome?. Gend Med 6: 152-167. https://doi.org/10.1016/j.genm.2009.03.004
|
| [80] |
Yung JA, Fuseini H, Newcomb DC (2018) Hormones, sex, and asthma. Ann Allerg Asthma Im 120: 488-494. https://doi.org/10.1016/j.anai.2018.01.016
|
| [81] |
Hall JM, Couse JF, Korach KS (2001) The multifaceted mechanisms of estradiol and estrogen receptor signaling. J Biol Chem 276: 36869-36872. https://doi.org/10.1074/jbc.R100029200
|
| [82] |
Yaşar P, Ayaz G, User SD, et al. (2017) Molecular mechanism of estrogen–estrogen receptor signaling. Reprod Med Biol 16: 4-20. https://doi.org/10.1002/rmb2.12006
|
| [83] |
Kovats S (2015) Estrogen receptors regulate innate immune cells and signaling pathways. Cell Immunol 294: 63-69. https://doi.org/10.1016/j.cellimm.2015.01.018
|
| [84] |
Cunningham M, Gilkeson G (2011) Estrogen receptors in immunity and autoimmunity. Clin Rev Allerg Immu 40: 66-73. https://doi.org/10.1007/s12016-010-8203-5
|
| [85] |
Bonds RS, Midoro-Horiuti T (2013) Estrogen effects in allergy and asthma. Curr Opin Allergy Clin Immunol 13: 92-99. https://doi.org/10.1097/ACI.0b013e32835a6dd6
|
| [86] |
Watanabe Y, Tajiki-Nishino R, Tajima H, et al. (2019) Role of estrogen receptors α and β in the development of allergic airway inflammation in mice: A possible involvement of interleukin 33 and eosinophils. Toxicology 411: 93-100. https://doi.org/10.1016/j.tox.2018.11.002
|
| [87] |
Graham JH, Yoachim SD, Gould KA (2020) Estrogen receptor alpha signaling is responsible for the female sex bias in the loss of tolerance and immune cell activation induced by the lupus susceptibility locus Sle1b. Front Immunol 11: 582214. https://doi.org/10.3389/fimmu.2020.582214
|
| [88] |
Gandhi VD, Cephus JY, Norlander AE, et al. (2022) Androgen receptor signaling promotes Treg suppressive function during allergic airway inflammation. J Clin Invest 132: e153397. https://doi.org/10.1172/JCI153397
|
| [89] |
Cephus JY, Stier MT, Fuseini H, et al. (2017) Testosterone attenuates group 2 innate lymphoid cell-mediated airway inflammation. Cell Rep 21: 2487-2499. https://doi.org/10.1016/j.celrep.2017.10.110
|
| [90] |
Laffont S, Blanquart E, Savignac M, et al. (2017) Androgen signaling negatively controls group 2 innate lymphoid cells. J Exp Med 214: 1581-1592. https://doi.org/10.1084/jem.20161807
|
| [91] |
Markle JG, Fish EN (2014) SeXX matters in immunity. Trends Immunol 35: 97-104. https://doi.org/10.1016/j.it.2013.10.006
|
| [92] |
Kissick HT, Sanda MG, Dunn LK, et al. (2014) Androgens alter T-cell immunity by inhibiting T-helper 1 differentiation. P Natl Acad Sci USA 111: 9887-9892. https://doi.org/10.1073/pnas.1402468111
|
| [93] |
Trigunaite A, Dimo J, Jørgensen TN (2015) Suppressive effects of androgens on the immune system. Cell Immunol 294: 87-94. https://doi.org/10.1016/j.cellimm.2015.02.004
|
| [94] |
García-Gómez E, González-Pedrajo B, Camacho-Arroyo I (2013) Role of sex steroid hormones in bacterial–host interactions. Biomed Res Int 2013: 928290. https://doi.org/10.1155/2013/928290
|
| [95] |
Meier A, Chang JJ, Chan ES, et al. (2009) Sex differences in the Toll-like receptor-mediated response of plasmacytoid dendritic cells to HIV-1. Nat Med 15: 955-959. https://doi.org/10.1038/nm.2004
|
| [96] | Afshan G, Afzal N, Qureshi S (2012) CD4+CD25(hi) regulatory T cells in healthy males and females mediate gender difference in the prevalence of autoimmune diseases. Clin Lab 58: 567-571. |
| [97] |
Fuseini H, Yung JA, Cephus JY, et al. (2018) Testosterone decreases house dust mite-induced type 2 and IL-17A-mediated airway inflammation. J Immunol 201: 1843-1854. https://doi.org/10.4049/jimmunol.1800293
|
| [98] |
Marozkina N, Zein J, DeBoer MD, et al. (2019) Dehydroepiandrosterone supplementation may benefit women with asthma who have low androgen levels: a pilot study. Pulm Ther 5: 213-220. https://doi.org/10.1007/s41030-019-00101-9
|
| [99] |
Zein J, Gaston B, Bazeley P, et al. (2020) HSD3B1 genotype identifies glucocorticoid responsiveness in severe asthma. P Natl Acad Sci USA 117: 2187-2193. https://doi.org/10.1073/pnas.1918819117
|
| [100] |
Wenzel SE, Robinson CB, Leonard JM, et al. (2010) Nebulized dehydroepiandrosterone-3-sulfate improves asthma control in the moderate-to-severe asthma results of a 6-week, randomized, double-blind, placebo-controlled study. Allergy Asthma Proc 31: 461-471. https://doi.org/10.2500/aap.2010.31.3384
|
| [101] |
Borba VV, Zandman-Goddard G, Shoenfeld Y (2018) Prolactin and autoimmunity. Front Immunol 9: 73. https://doi.org/10.3389/fimmu.2018.00073
|
| [102] |
Moulton VR (2018) Sex hormones in acquired immunity and autoimmune disease. Front Immunol 9: 2279. https://doi.org/10.3389/fimmu.2018.02279
|
| [103] |
Ngo ST, Steyn FJ, McCombe PA (2014) Gender differences in autoimmune disease. Front Neuroendocrin 35: 347-369. https://doi.org/10.1016/j.yfrne.2014.04.004
|
| [104] |
Keselman A, Fang X, White PB, et al. (2017) Estrogen signaling contributes to sex differences in macrophage polarization during asthma. J Immunol 199: 1573-1583. https://doi.org/10.4049/jimmunol.1601975
|
Figures(1) / Tables(1)

McKenna S. Vininski, Sunanda Rajput, Nicholas J. Hobbs, Joseph J. Dolence. Understanding sex differences in the allergic immune response to food[J]. AIMS Allergy and Immunology, 2022, 6(3): 90-105. doi: 10.3934/Allergy.2022009







 DownLoad:
DownLoad: