Citation: Larisa I. Fedoreyeva, Boris F. Vanyushin. Possible existence of a system similar to bacterial restriction–modification in plants[J]. AIMS Molecular Science, 2020, 7(4): 396-413. doi: 10.3934/molsci.2020020

| [1] |
Rao DN, Dryden DTF, Bheemanaik S (2014) Type III restriction-modification enzymes: a historical perspective. Nucleic Acids Res 42: 45-55. doi: 10.1093/nar/gkt616

|
| [2] |
Loenen WAM, Raleigh EA (2014) The other face of restriction: modification-dependent enzymes. Nucleic Acids Res 42: 56-69. doi: 10.1093/nar/gkt747
|
| [3] |
Dixon M, Fauman EB, Ludwig ML (1999) The black sheep of the family: AdoMet-dependent methyltransferases that do not fit the consensus structural fold. S-Adenosylmethionine-dependent Methyltransferases: Structures and Functions World Scientific Inc. Singapore: 39-54. doi: 10.1142/9789812813077_0002
|
| [4] |
Wilson GG, Murray NE (1991) Restriction and modification systems. Annu Rev Genet 5: 585-627. doi: 10.1146/annurev.ge.25.120191.003101
|
| [5] |
Arber W, Dussoix D (1962) Host specificity of DNA produced by Escherichia coli. I. Host controlled modification of bacteriophage lambda. J Mol Biol 5: 18-36. doi: 10.1016/S0022-2836(62)80058-8
|
| [6] |
Roberts RJ, Belfort M, Bestor T, et al. (2003) A nomenclature for restriction enzymes, DNA methyltransferases, homing endonucleases and their genes. Nucleic Acids Res 31: 1805-1812. doi: 10.1093/nar/gkg274
|
| [7] | Marinus MGMethylation of DNA. Escherichia coli and Salmonella typhimurium (1996) .Washington DC: ASM Press, 782-791. |
| [8] |
Peterson KR, Wertman KF, Mount DW, et al. (1985) Viability of Escherichia coli K-12 DNA adenine methylase (dam) mutants requires increased expression of specific genes in the SOS regulon. Mol Gen Gene 201: 14-19. doi: 10.1007/BF00397979
|
| [9] |
Lieb M, Bhagwat AS (1996) Very short patch repair: Reducing the cost of cytosine methylation. Mol Microbiol 20: 467-473. doi: 10.1046/j.1365-2958.1996.5291066.x
|
| [10] |
Ban C, Yang W (1998) Structural basis for MutH activation in E. coli mismatch repair and relationship of MutH to restriction endonucleases. EMBO J 17: 1526-1534. doi: 10.1093/emboj/17.5.1526
|
| [11] |
Ahmad I, Rao DN (1996) Chemistry and biology of DNA methyltransferases. Crit Rev Biochem Mol Biol 31: 361-380. doi: 10.3109/10409239609108722
|
| [12] |
Gong W, O'Gara M, Blumenthal RM, et al. (1997) Structure of PvuII DNA-(cytosine N4) methyltransferase, an example of domain permutation and protein fold assignment. Nucleic Acids Res 25: 2702-2715. doi: 10.1093/nar/25.14.2702
|
| [13] |
Stoddard BL (2005) Homing endonuclease structure and function. Q Rev Biophys 38: 49-95. doi: 10.1017/S0033583505004063
|
| [14] |
Spiegel PC, Chevalier B, Sussman D, et al. (2006) The structure of I-CeuI homing endonuclease: Evolving asymmetric DNA recognition from a symmetric protein scaffold. Structure 14: 869-880. doi: 10.1016/j.str.2006.03.009
|
| [15] | Blow MJ, Clark TA, Daum CG, et al. (2016) The Epigenomic Landscape of Prokaryotes. PLoS Genet 12. |
| [16] |
Vasu K, Nagaraja V (2013) Diverse functions of restriction-modification systems in addition to cellular defense. Microbiol Mol Biol Rev 77: 53-72. doi: 10.1128/MMBR.00044-12
|
| [17] | Vanyushin BF, Ashapkin VV (2009) DNA Methylation in Plants New York: Nova Science Publishers Inc. |
| [18] | Vanyushin BF, Ashapkin VV (2007) Peculiarities of DNA methylation in plants. Progress in DNA Methylation Research New York: Nova Science Publishers Inc., 1-90. |
| [19] |
Goll MG, Bestor TH (2005) Eukaryotic cytosine methyltransferases. Annu Rev Biochem 74: 481-514. doi: 10.1146/annurev.biochem.74.010904.153721
|
| [20] |
Lister R, Pelizzola M, Dowen RH, et al. (2009) Human DNA methylomes at base resolution show widespread epigenomic differences. Nature 462: 315-322. doi: 10.1038/nature08514
|
| [21] |
Lister R, Mukamel EA, Nery JR, et al. (2013) Global epigenomic reconfiguration during mammalian brain development. Science 341: 1237905. doi: 10.1126/science.1237905
|
| [22] |
Greer EL, Blanco MA, Gu L, et al. (2015) DNA Methylation on N6-Adenine in C. elegans. Cell 7: 868-878. doi: 10.1016/j.cell.2015.04.005
|
| [23] |
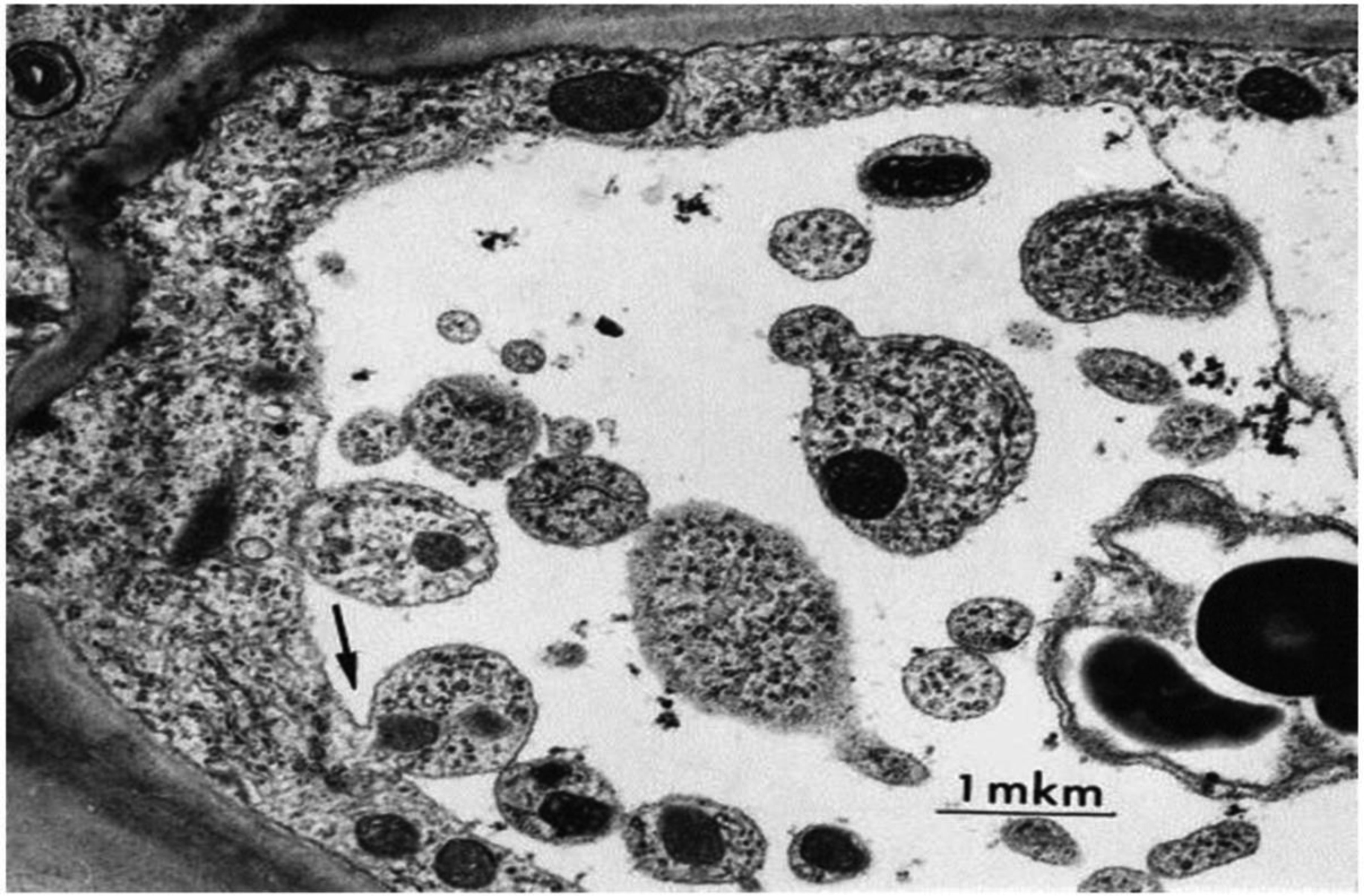
Bakeeva LE, Kirnos MD, Aleksandrushkina NI, et al. (1999) Subcellular reorganization of mitochondria producing heavy DNA in aging wheat coleoptiles. FEBS Lett 457: 122-125. doi: 10.1016/S0014-5793(99)01025-X
|
| [24] |
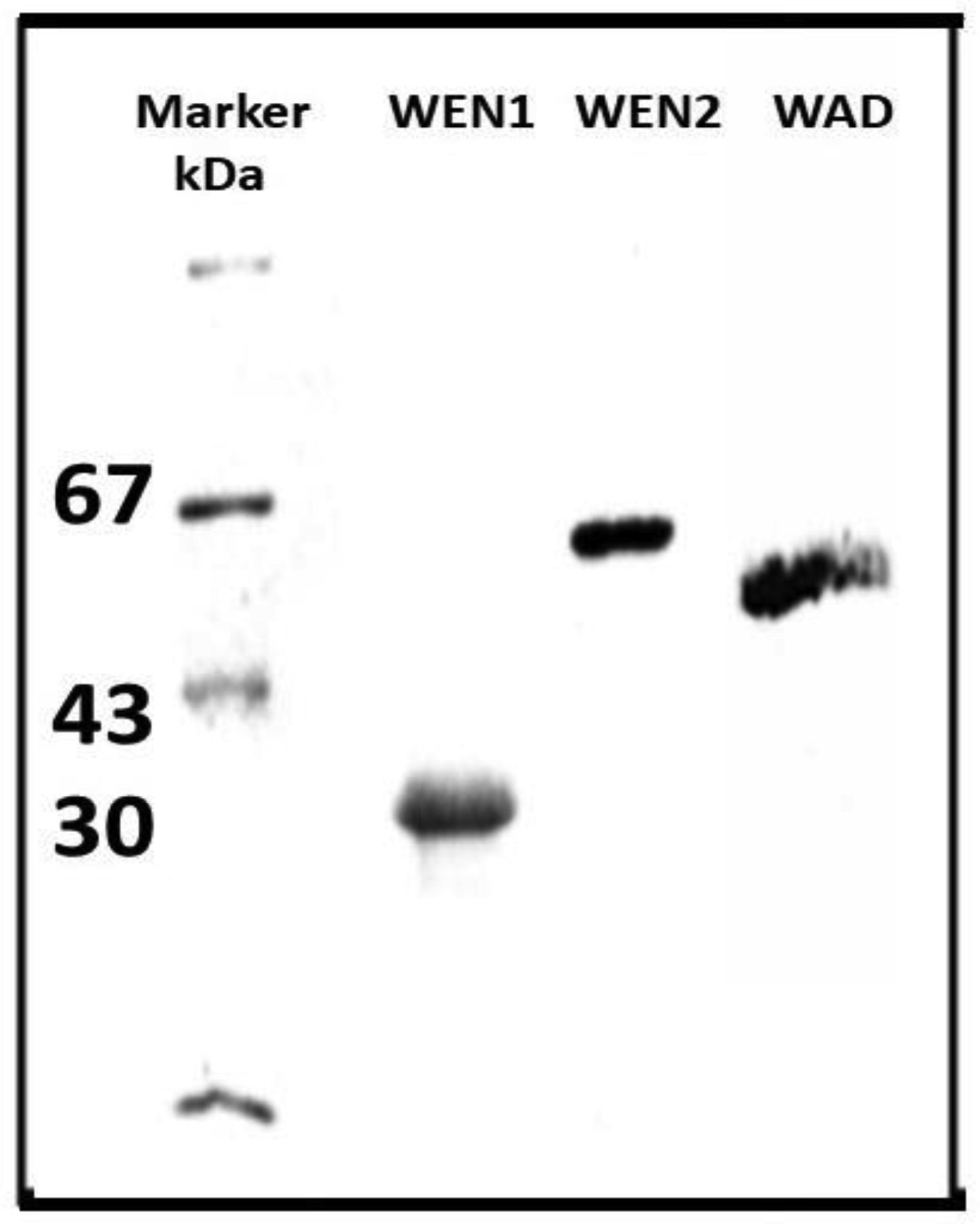
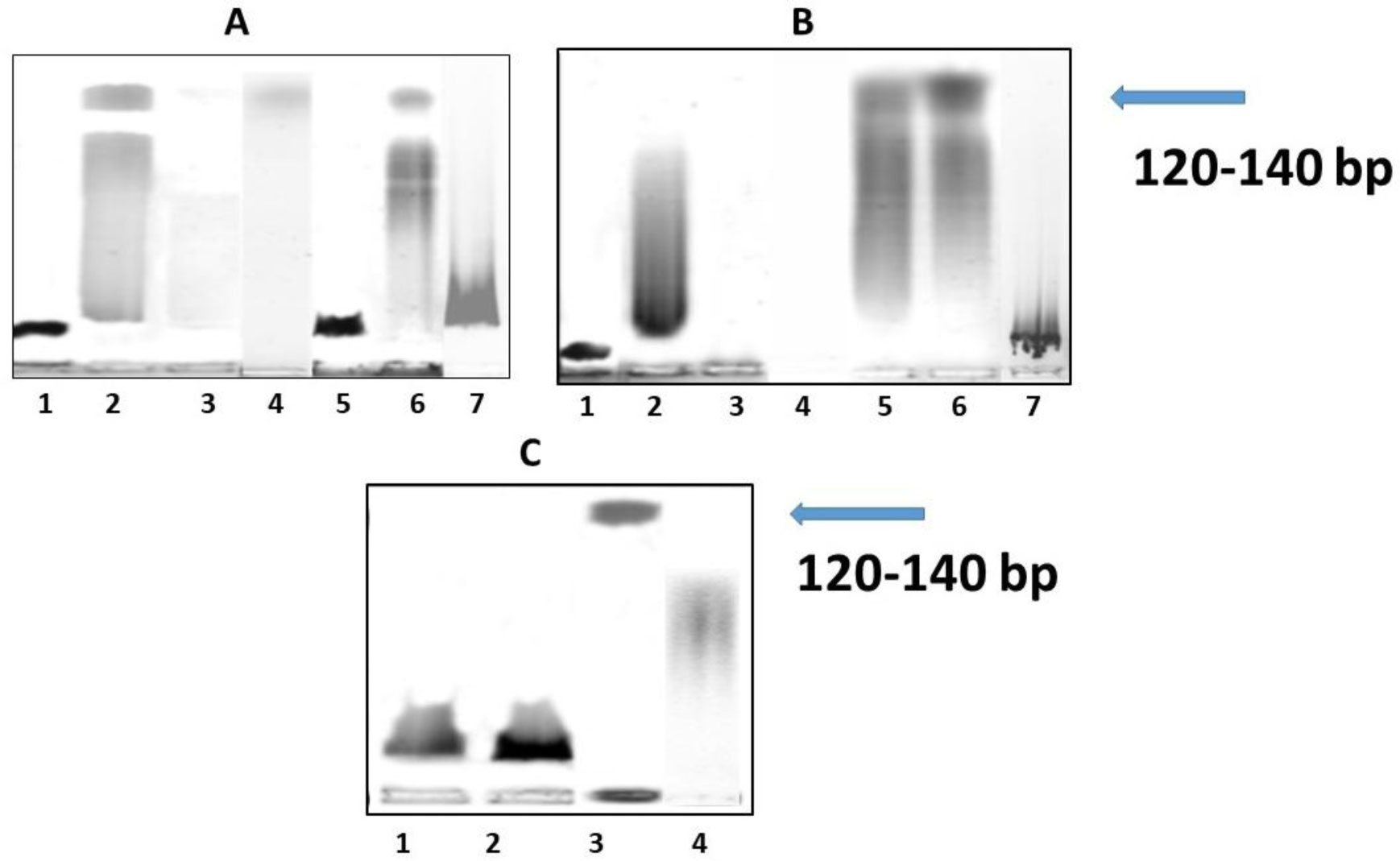
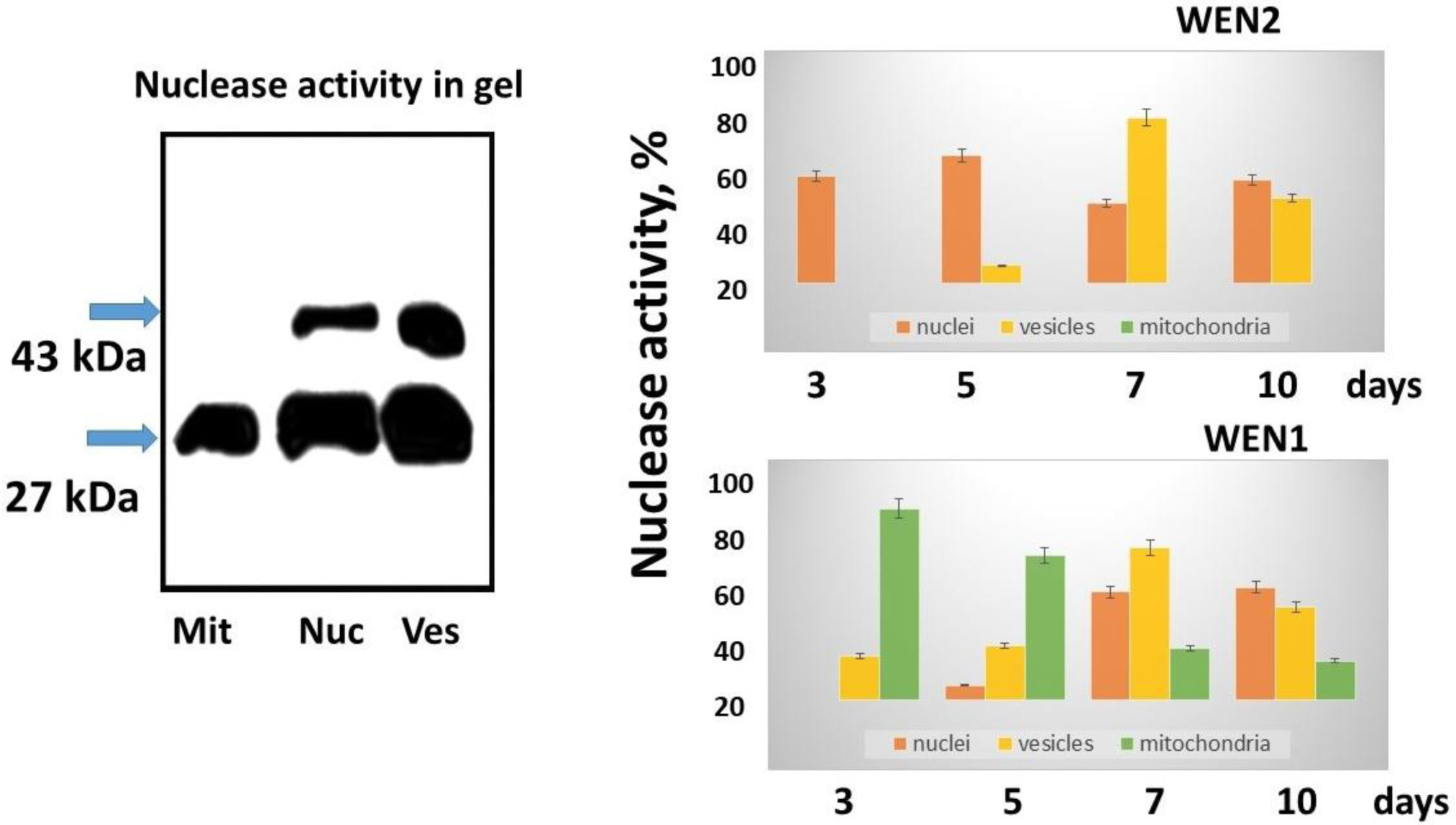
Fedoreyeva LI, Sobolev DE, Vanyushin BF (2007) Wheat endonuclease WEN1 depent on S-adenosyl-L-methionine and sensitive to DNA methylation status. Epigenetics 2: 50-53. doi: 10.4161/epi.2.1.3933
|
| [25] | Fedoreyeva LI, Sobolev DE, Vanyushin BF (2008) S-adenosyl-L-methionine dependent and sensitive to the status of DNA methylation endonuclease WEN2 from wheat coleoptiles. Biochemistry (Russian) 73: 1243-1251. |
| [26] |
Fedoreyeva LI, Vanyushin B F (2002) N6-Adenine DNA-methyltransferase in wheat seedlings. FEBS Lett 514: 305-308. doi: 10.1016/S0014-5793(02)02384-0
|
| [27] | Kirnos MD, Aleksandrushkina NI, Vanyushin BF (1997) Apoptosis in cells of the first leaf and coleoptile of wheat seedlings: internucleosomal fragmentation of the genome and synthesis of heavy oligonucleosomal size of DNA fragments. Biochemistry (Russian) 62: 1008-1014. |
| [28] |
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analyt Biochem 72: 248-254. doi: 10.1016/0003-2697(76)90527-3
|
| [29] |
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685. doi: 10.1038/227680a0
|
| [30] |
Bannister D, Glover SW (1968) Restriction and modification of bacteriophages by R strains of Escherichia coli K-12. Biochem Biophys Res Commun 30: 735-738. doi: 10.1016/0006-291X(68)90575-5
|
| [31] |
Vertino PM (1999) Eukaryotic DNA methyltransferases. S-Adenosylmethionine-dependent Methyltransferases: Structures and Functions 341-372. doi: 10.1142/9789812813077_0012
|
| [32] |
Bist P, Sistla S, Krishnamurthy V, et al. (2001) S-Adenosyl-L-methionine is required for DNA cleavage by type III restriction enzymes. J Mol Biol 310: 93-109. doi: 10.1006/jmbi.2001.4744
|
| [33] |
Cowan JA (2002) Structural and catalytic chemistry of magnesium-dependent enzymes. Biometals 15: 225-235. doi: 10.1023/A:1016022730880
|
| [34] | Lyons TJ, Eide DJ (2006) Transport and storage of metal ions in biology. Biological Inorganic Chemistry: Structure and Reactivity 57-77. |
| [35] |
Maguire ME, Cowan JA (2002) Magnesium chemistry and biochemistry. Biometals 15: 203-210. doi: 10.1023/A:1016058229972
|
| [36] |
Romani A, Scarpa A (1992) Regulation of cell magnesium. Arch Biochem Biophys 298: 1-12. doi: 10.1016/0003-9861(92)90086-C
|
| [37] | Kirnos MD, Aleksandrushkina NI, Goremykin VV, et al. (1992) “Heavy” mitochondrial DNA in higher plants. Biochemistry (Russian) 57: 1566-1673. |
| [38] | Aleksandrushkina NI, Kudryashova IB, Kirnos MD, et al. (1990) Synthesis and methylation at adenine residues of “heavy” miniplasmids of mitochondrial DNA in coleoptile and leaves of wheat seedlings. The influence of phytohormones. Biochemistry (Russian) 55: 2038-2045. |
| [39] |
Kirnos MD, Alexandrushkina NI, Zagorskaya GYa, et al. (1992) Superproduction of heavy minicircular mitochondrial DNA in aging wheat coleoptiles. FEBS Lett 298: 109-112. doi: 10.1016/0014-5793(92)80033-D
|

Figures(9) / Tables(1)

Larisa I. Fedoreyeva, Boris F. Vanyushin. Possible existence of a system similar to bacterial restriction–modification in plants[J]. AIMS Molecular Science, 2020, 7(4): 396-413. doi: 10.3934/molsci.2020020







 DownLoad:
DownLoad: