Citation: Kenji Chiba, Yasuhiro Maeda, Noriyasu Seki, Hirotoshi Kataoka, Kunio Sugahara. Role of sphingosine 1-phosphate (S1P) and effects of fingolimod, an S1P receptor 1 functional antagonist in lymphocyte circulation and autoimmune diseases[J]. AIMS Molecular Science, 2014, 1(4): 162-182. doi: 10.3934/molsci.2014.4.162

| [1] | Fujita T, Inoue K, Yamamoto S, et al. (1994) Fungal metabolites. Part 11. A potent immunosuppressive activity found in Isaria sinclairii metabolite. J Antibiot 47: 208-215. |
| [2] | Fujita T, Inoue K, Yamamoto S, et al. (1994) Fungal metabolites. Part 12. Potent immunosuppressant, 14-deoxomyruiocin, (2S,3R,4R)-(E)-2-amino-3,4-dihydroxy-2- hydroxy- methyleicos-6-enoic acid and structure-activity relationships of myriocin derivatives. J Antibiot 47: 216-224. |
| [3] |
Fujita T, Yoneta M, Hirose R, et al. (1995) Simple compounds, 2-alkyl-2- amino-1,3-propane-diols have potent immunosuppressive activity. Bioorg Med Chem Lett 5: 847-852. doi: 10.1016/0960-894X(95)00126-E

|
| [4] |
Adachi K, Kohara T, Nakao N, et al. (1995) Design, synthesis, and structure-activity relationships of 2-substituted-2-amino-1,3-propanediols: Discovery of a novel immuno- suppressant, FTY720. Bioorg Med Chem Lett 5: 853-856. doi: 10.1016/0960-894X(95)00127-F
|
| [5] |
Fujita T, Hirose R, Yoneta M, et al. (1996) Potent immunosuppressants, 2-alkyl-2- aminopropane-1,3-diols. J Med Chem 39: 4451-4459. doi: 10.1021/jm960391l
|
| [6] |
Kiuchi M, Adachi K, Kohara T, et al. (2000) Synthesis and immunosuppressive activity of 2-substituted 2-aminopropane-1,3-diols and 2-aminoethanols. J Med Chem 43: 2946-2961. doi: 10.1021/jm000173z
|
| [7] | Adachi K, Chiba K (2008) FTY720 story. Its discovery and the following accelerated development of sphingosine 1-phosphate receptor agonists as immunomodulators based on reverse pharmacology. Perspect Medicin Chem 1: 11-23. |
| [8] |
Chiba K, Adachi K (2012) Sphingosine 1-phosphate receptor 1 as a useful target for treatment of multiple sclerosis. Pharmaceuticals 5:514-528. doi: 10.3390/ph5050514
|
| [9] | Chiba K, Hoshino Y, Suzuki C, et al. (1996) FTY720, a novel immunosuppressant possessing unique mechanisms. I. Prolongation of skin allograft survival and synergistic effect in combination with cyclosporine in rats. Transplant Proc 28: 1056-1059. |
| [10] | Hoshino Y, Suzuki C, Ohtsuki M, et al. (1996) FTY720, a novel immunosuppressant possessing unique mechanisms. II. Long-term graft survival induction in rat heterotopic cardiac allografts and synergistic effect in combination with cyclosporine A. Transplant Proc 28: 1060-1061. |
| [11] | Kawaguchi T, Hoshino Y, Rahman F, et al. (1996) FTY720, a novel immunosuppressant possessing unique mechanisms. III. Synergistic prolongation of canine renal allograft survival in combination with cyclosporine A. Transplant Proc 28: 1062-1063. |
| [12] |
Chiba K. (2005) FTY720, a new class of immunomodulator, inhibits lymphocyte egress from secondary lymphoid tissues and thymus by agonistic activity at sphingosine 1-phosphate receptors. Pharmacol Ther 108: 308-319. doi: 10.1016/j.pharmthera.2005.05.002
|
| [13] |
Matsuura M, Imayoshi T, Chiba K, Okumoto T (2000) Effect of FTY720, a novel immunosuppressant, on adjuvant-induced arthritis in rats. Inflamm Res 49: 404-410. doi: 10.1007/s000110050608
|
| [14] | Chiba K, Yanagawa Y, Masubuchi Y, et al. (1998) FTY720, a novel immunosuppressant, induces sequestration of circulating mature lymphocytes by acceleration of lymphocyte homing in rats. I. FTY720 selectively decreases the number of circulating mature lymphocytes by acceleration of lymphocyte homing. J Immunol 160: 5037-5044. |
| [15] | Yanagawa Y, Sugahara K, Kataoka H, et al. (1998) FTY720, a novel immunosuppressant, induces sequestration of circulating mature lymphocytes by acceleration of lymphocyte homing in rats. II. FTY720 prolongs skin allograft survival by decreasing T cell infiltration into grafts but not cytokine production in vivo. J Immunol 160: 5493-5499. |
| [16] |
Brinkmann V. Pinschewer D, Chiba K, Feng L (2000) FTY720: A novel transplantation drug that modulates lymphocyte traffic rather than activation. Trends Pharmacol Sci 21: 49-52. doi: 10.1016/S0165-6147(99)01419-4
|
| [17] |
Brinkmann V, Davis MD, Heise CE, et al. (2002) The immune modulator FTY720 targets sphingosine 1-phosphate receptors. J Biol Chem 277: 21453-21457. doi: 10.1074/jbc.C200176200
|
| [18] |
Mandala S, Hajdu R, Bergstrom J, et al. (2002) Alteration of lymphocyte trafficking by sphingosine-1-phosphate receptor agonists. Science 296: 346-349. doi: 10.1126/science.1070238
|
| [19] |
Matloubian M, Lo CG, Cinamon G, et al. (2004) Lymphocyte egress from thymus and peripheral lymphoid organs is dependent on S1P receptor 1. Nature 427: 355-360. doi: 10.1038/nature02284
|
| [20] |
Spiegel S, Milstien S (2007) Functions of the multifaceted family od sphingosine kinases and some dose relatives. J Biol Chem 282: 2125-2129. doi: 10.1074/jbc.R600028200
|
| [21] |
Hannun YA, Obeid LM (2008) Principles of bioactive lipid signalling: lessons from sphingolipids. Nat Rev Mol Cell Biol 9: 139-150. doi: 10.1038/nrm2329
|
| [22] |
Pyne S, Pyne N (2000) Sphingosine 1-phosphate signalling via the endothelial differentiation gene family of G-protein-coupled receptors. Pharmacol Ther 88: 115-131. doi: 10.1016/S0163-7258(00)00084-X
|
| [23] | Spiegel S (2000) Sphingosine 1-phosphate: A ligand for the EDG-1 family of G-protein-coupled receptors. Ann NY Acad Sci 905: 54-60. |
| [24] |
Rivera J, Proia RL, Olivera A (2008) The alliance of shpingosine-1-phosphate and its receptors in immunity. Nat Rev Immunol 8: 753-763. doi: 10.1038/nri2400
|
| [25] |
Spiegel S, Milstien S (2011) The outs and the ins of shingosine-1-phosphate in immunity. Nat Rev Immunol 11: 403-415. doi: 10.1038/nri2974
|
| [26] |
Paugh SW, Payne SG, Barbour SE, et al. (2003) The immunosuppressant FTY720 is phosphorylated by sphingosine kinase type 2. FEBS Lett 554:189-193. doi: 10.1016/S0014-5793(03)01168-2
|
| [27] | Chiba K, Matsuyuki H, Maeda Y, Sugahara K (2006) Role of sphingosine 1-phosphate receptor type 1 in lymphocyte egress from secondary lymphoid tissues and thymus. Cell Mol Immunol 3: 11-19. |
| [28] |
Maeda Y, Matsuyuki H, Shimano K, et al. (2007) Migration of CD4 T cells and dendritic cells toward sphingosine 1-phosphate (S1P) is mediated by different receptor subtypes: S1P regulates the functions of murine mature dendritic cells via S1P receptor type 3. J Immunol 178: 3437-3446. doi: 10.4049/jimmunol.178.6.3437
|
| [29] |
Pham TH, Okada T, Matloubian M, et al. (2008) S1P1 receptor signaling overrides retention mediated by G alpha i-coupled receptors to promote T cell egress. Immunity 28: 122-133. doi: 10.1016/j.immuni.2007.11.017
|
| [30] |
Cyster JG (2005) Chemokines, sphingosine-1-phosphate, and cell migration in secondary lymphoid organs. Annu Rev Immunol 23: 127-159. doi: 10.1146/annurev.immunol.23.021704.115628
|
| [31] |
Mitra P, Oskeritzian CA, Payne SG, et al. (2006) Role of ABCC1 in export of sphingosine- 1-phosphate from mast cells. Proc Natl Acad Sci USA 103: 16394-16399. doi: 10.1073/pnas.0603734103
|
| [32] |
Nishi T, Kobayashi N, Hisano Y et al. (2014) Molecular and physiological functions of sphingosine 1-phosphate transporters. Biochim Biophys Acta 1841: 759-765. doi: 10.1016/j.bbalip.2013.07.012
|
| [33] |
Mechtcheriakova D, Wlachos A, Sobanov J, et al. (2007) Sphingosine 1-phosphate phosphatase 2 is induced during inflammatory responses. Cell Signal 19: 748-760. doi: 10.1016/j.cellsig.2006.09.004
|
| [34] |
Peest U, Sensken SC, Andréani P, et al. (2008) S1P-lyase independent clearance of extracellular sphingosine 1-phosphate after dephosphorylation and cellular uptake. J Cell Biochem 104: 756-772. doi: 10.1002/jcb.21665
|
| [35] |
Zhao Y, Kalari SK, Usatyuk PV, et al. (2007) Intracellular generation of sphingosine 1-phosphate in human lung endothelial cells: role of lipid phosphate phosphatase-1 and sphingosine kinase 1. J Biol Chem 282: 14165-14177. doi: 10.1074/jbc.M701279200
|
| [36] |
Pappu R, Schwab SR, Cornelissen I, et al. (2007) Promotion of lymphocyte egress into blood and lymph by distinct sources of sphingosine-1-phosphate. Science 316: 295-298. doi: 10.1126/science.1139221
|
| [37] |
Schwab SR, Pereira JP, Matloubian M, et al. (2005) Lymphocyte sequestration through S1P lyase inhibition and disruption of S1P gradients. Science 309: 1735-1739. doi: 10.1126/science.1113640
|
| [38] |
Lo CG, Xu Y, Proia RL, Cyster JG (2005) Cyclical modulation of sphingosine-1-phosphate receptor 1 surface expression during lymphocyte recirculation and relationship to lymphoid organ transit. J Exp Med. 201: 291-301. doi: 10.1084/jem.20041509
|
| [39] |
Kiuchi M, Adachi K, Tomatsu A, et al. (2005) Asymmetric synthesis and biological evaluation of the enantiomeric isomers of the immunosuppressive FTY720-phosphate. Bioorg. Med. Chem 13: 425-432. doi: 10.1016/j.bmc.2004.10.008
|
| [40] | Matsuyuki H, Maeda Y, Yano K, et al. (2006) Involvement of sphingosine 1-phosphate (S1P) receptor type 1 and type 4 in migratory response of mouse T cells toward S1P. Cell Mol Immunol 3: 429-437. |
| [41] |
Oo ML, Thangada S, Wu MT, et al. (2007) Immunosuppressive and anti-angiogenic sphingosine 1-phosphate receptor-1 agonists induce ubiquitinylation and proteasomal degradation of the receptor. J Biol Chem 282: 9082-9089. doi: 10.1074/jbc.M610318200
|
| [42] |
Ladi E, Yin X, Chtanova T, Robey EA (2006) Thymic microenvironments for T cell differentiation and selection. Nat Immunol 7: 338-343. doi: 10.1038/ni1323
|
| [43] |
Drennan MB, Elewaut D, Hogquist KA (2009) Thymic emigration: sphingosine-1-phosphate receptor-1-dependent models and beyond. Eur J Immunol 39: 925-930. doi: 10.1002/eji.200838912
|
| [44] | Kato S, Schoefl GI (1989) Microvasculature of normal and involuted mouse thymus. Light- and electron-microscopic study. Acta Anat (Basel) 135: 1-11. |
| [45] |
Kato S (1997) Thymic microvascular system. Microsc Res Tech 38: 287-299. doi: 10.1002/(SICI)1097-0029(19970801)38:3<287::AID-JEMT9>3.0.CO;2-J
|
| [46] |
Mori K, Itoi M, Tsukamoto N, et al. (2007) The perivascular space as a path of hematopoietic progenitor cells and mature T cells between the blood circulation and the thymic parenchyma. Int Immunol 19: 745-753. doi: 10.1093/intimm/dxm041
|
| [47] |
Zachariah MA, Cyster JG (2010) Neural crest-derived pericytes promote egress of mature thymocytes at the corticomedullary junction. Science 328: 1129-1135. doi: 10.1126/science.1188222
|
| [48] |
Cyster JG, Schwab SR (2012) Sphingosine-1-phosphate and lymphocyte egress from lymphoid organs. Annu Rev Immunol 30: 69-94. doi: 10.1146/annurev-immunol-020711-075011
|
| [49] | Yagi H, Kamba R, Chiba K, et al. (2000) Immunosuppressant FTY720 inhibits thymocyte emigration. Eur J Immunol 30:1435. |
| [50] |
Maeda Y, Yagi H, Takemoto K, et al. (2014) S1P lyase in thymic perivascular space promotes egress of mature thymocytes via up-regulation of S1P receptor 1. Int Immunol 26: 245-255. doi: 10.1093/intimm/dxt069
|
| [51] |
Allende ML, Dreier JL, Mandala S, Proia RL (2004) Expression of the sphingosine 1-phosphate receptor, S1P1, on T-cells controls thymic emigration. J Biol Chem 279: 15396-15401. doi: 10.1074/jbc.M314291200
|
| [52] |
Saba JD, Hla T (2004) Point-counterpoint of sphingosine 1-phosphate metabolism. Circ Res 94: 724-734. doi: 10.1161/01.RES.0000122383.60368.24
|
| [53] |
Breart B, Ramos-Perez WD, Mendoza A, et al. (2011) Lipid phosphate phosphatase 3 enables efficient thymic egress. J Exp Med 208: 1267-1278. doi: 10.1084/jem.20102551
|
| [54] | Vogel P, Donoviel MS, Read R, et al. (2009) Incomplete inhibition of sphingosine 1-phosphate lyase modulates immune system function yet prevents early lethality and non-lymphoid lesions. PLoS One 4:e4112. |
| [55] |
Borowsky AD, Bandhuvula P, Kumar A, , et al. (2012) Sphingosine-1-phosphate lyase expression in embryonic and adult murine tissues. J Lipid Res 53: 1920-1931. doi: 10.1194/jlr.M028084
|
| [56] |
Suzuki S, Li XK, Enosawa S, and Shinomiya T (1996) A new immunosuppressant, FTY720, induces bcl-2-associated apoptotic cell death in human lymphocytes. Immunology 89:518-23. doi: 10.1046/j.1365-2567.1996.d01-777.x
|
| [57] |
Nagahara Y, Enosawa S, Ikekita M, et al. (2000) Evidence that FTY720 induces T cell apoptosis in vivo. Immunopharmacology 48:75-85. doi: 10.1016/S0162-3109(00)00181-8
|
| [58] |
Luo ZJ, Tanaka T, Kimura F, Miyasaka M (1999) Analysis of the mode of action of a novel immunosuppressant FTY720 in mice. Immunopharmacology. 41:199-207. doi: 10.1016/S0162-3109(99)00004-1
|
| [59] |
Sugito K, Koshinaga T, Inoue M, et al. (2005) The effect of a novel immunosuppressant, FTY720, in mice without secondary lymphoid organs. Surg Today 35:662-7. doi: 10.1007/s00595-005-3011-x
|
| [60] |
Maeda Y, Seki N, Sato N, et al. (2010) Sphingosine 1-phosphate receptor type 1 regulates egress of mature T cells from mouse bone marrow. Int Immunol 22: 515-525. doi: 10.1093/intimm/dxq036
|
| [61] |
Webb M, Tham CS, Lin FF, et al. (2004) Sphingosine 1-phosphate receptor agonists attenuate relapsing-remitting experimental autoimmune encephalitis in SJL mice. J Neuroimmunol 153: 108-121. doi: 10.1016/j.jneuroim.2004.04.015
|
| [62] | Kataoka H, Sugahara K, Shimano K, et al. (2005) FTY720, sphingosine 1-phosphate receptor modulator, ameliorates experimental autoimmune encephalomyelitis by inhibition of T cell infiltration. Cell Mol Immunol 2: 439-448. |
| [63] |
Balatoni B, Storch MK, Swoboda EM, et al. (2007) FTY720 sustains and restores neuronal function in the DA rat model of MOG-induced experimental autoimmune encephalomyelitis. Brain Res Bull 74: 307-316. doi: 10.1016/j.brainresbull.2007.06.023
|
| [64] |
Foster CA, Howard LM, Schweitzer A, et al. (2007) Brain penetration of the oral immunomodulatory drug FTY720 and its phosphorylation in the central nervous system during experimental autoimmune encephalomyelitis: Consequences for mode of action in multiple sclerosis. J Pharmacol Exp Ther 323: 469-475. doi: 10.1124/jpet.107.127183
|
| [65] |
Brinkmann V (2007) Sphingosine 1-phosphate receptors in health and disease: Mechanistic insights from gene deletion studies and reverse pharmacology. Pharmacol Ther 115: 84-105. doi: 10.1016/j.pharmthera.2007.04.006
|
| [66] |
Chiba K, Kataoka H, Seki N, et al. (2011) Fingolimod (FTY720), sphingosine 1-phosphate receptor modulator, shows superior efficacy as compared with interferon-β in mouse experimental autoimmune encephalomyelitis. Int Immunopharmacol 11: 366-372. doi: 10.1016/j.intimp.2010.10.005
|
| [67] |
Langrish CL, Chen Y, Blumenschein WM, et al. (2005) IL-23 drives a pathogenic T cell population that induces autoimmune inflammation. J Exp Med 201: 233-240. doi: 10.1084/jem.20041257
|
| [68] |
Komiyama Y, Nakae S, Matsuki T, et al. (2006) IL-17 plays an important role in the development of experimental autoimmune encephalomyelitis. J Immunol 177: 566-573. doi: 10.4049/jimmunol.177.1.566
|
| [69] |
Stromnes IM, Cerretti LM, Liggitt D, et al. (2008) Differential regulation of central nervous system autoimmunity by TH1 and TH17 cells. Nat Med 14: 337-342. doi: 10.1038/nm1715
|
| [70] | Seki N, Maeda Y, Kataoka H, et al. (2013) Role of sphingosine 1-phosphate (S1P) receptor 1 in experimental autoimmune encephalomyelitis. I. S1P-S1P1 axis induces migration of Th1 and Th17 cells. Pharmacology & Pharmacy 4: 628-637. |
| [71] |
Brinkmann V (2009) FTY720 (fingolimod) in Multiple Sclerosis: Therapeutic effects in the immune and the central nervous system. Br J Pharmacol 158: 1173-1182. doi: 10.1111/j.1476-5381.2009.00451.x
|
| [72] |
Choi JW, Gardell SE, Herr DR, et al. (2011) FTY720 (fingolimod) efficacy in an animal model of multiple sclerosis requires astrocyte sphingosine 1-phosphate receptor 1 (S1P1) modulation. Proc Natl Acad Sci USA 108: 751-756. doi: 10.1073/pnas.1014154108
|
| [73] | Seki N, Kataoka H, Sugahara K, et al. (2013) Role of sphingosine 1-phosphate (S1P) receptor 1 in experimental autoimmune encephalomyelitis. II. S1P-S1P1 axis induces pro-inflammatory cytokine production in astrocytes. Pharmacology & Pharmacy 4: 638-646. |
| [74] |
Matsuura M, Imayoshi T, Okumoto T (2000) Effect of FTY720, a novel immunosuppressant, on adjuvant- and collagen-induced arthritis in rats. Int J Immunopharmacol 22: 323-331. doi: 10.1016/S0192-0561(99)00088-0
|
| [75] |
Tsunemi S, Iwasaki T, Kitano S, et al. (2010) Effects of the novel immunosuppressant FTY720 in a murine rheumatoid arthritis model. Clin Immunol 136: 197-204. doi: 10.1016/j.clim.2010.03.428
|
| [76] | Okazaki H, Hirata D, Kamimura T, et al. (2002) Effects of FTY720 in MRL-lpr/lpr mice: therapeutic potential in systemic lupus erythematosus. J Rheumatol 29: 707-716. |
| [77] |
Wenderfer SE, Stepkowski SM, Braun MC (2008) Increased survival and reduced renal injury in MRL/lpr mice treated with a novel sphingosine-1-phosphate receptor agonist. Kidney Int 74: 1319-1326. doi: 10.1038/ki.2008.396
|
| [78] |
Alperovich G, Rama I, Lloberas N, et al. (2007) New immunosuppresor strategies in the treatment of murine lupus nephritis. Lupus 16: 18-24. doi: 10.1177/0961203306073136
|
| [79] |
Mizushima T, Ito T, Kishi D, et al. (2004) Therapeutic effects of a new lymphocyte homing reagent FTY720 in interleukin-10 gene-deficient mice with colitis. Inflamm Bowel Dis 10: 182-192. doi: 10.1097/00054725-200405000-00002
|
| [80] | Deguchi Y, Andoh A, Yagi Y, et al (2006) The S1P receptor modulator FTY720 prevents the development of experimental colitis in mice. Oncol Rep 16: 699-703. |
| [81] |
Daniel C, Sartory N, Zahn N, et al. (2007) FTY720 ameliorates Th1-mediated colitis in mice by directly affecting the functional activity of CD4+CD25+ regulatory T cells. J Immunol 178: 2458-2468. doi: 10.4049/jimmunol.178.4.2458
|
| [82] |
Radi ZA, Heuvelman DM, Masferrer JL, et al (2011) Pharmacologic evaluation of sulfasalazine, FTY720, and anti-IL-12/23p40 in a TNBS-induced Crohn's disease model. Dig Dis Sci 56: 2283-2291. doi: 10.1007/s10620-011-1628-8
|
| [83] | Budde K, Schmouder RL, Brunkhorst R, et al. (2002) Human first trail of FTY720, a novel immunomodulator, in stable renal transplant patients. J Am Soc Nephrol 13: 1073-1083. |
| [84] |
Budde K, Schmouder RL, Nashan B, et al. (2003) Pharmacodynamics of single doses of the novel immunosuppressant FTY720 in stable renal transplant patients. Am J Transplant 3: 846-854. doi: 10.1034/j.1600-6143.2003.00130.x
|
| [85] |
Kahan BD, Karlix JL, Ferguson RM, et al. (2003) Pharmacodynamics, pharmacokinetics, and safety of multiple doses of FTY720 in stable renal transplant patients: a multicenter, randomized, placebo-controlled, phase I study. Transplantation 76: 1079-1084. doi: 10.1097/01.TP.0000084822.01372.AC
|
| [86] |
Martin R, McFarland HF, McFarlin DE (1992) Immunological aspects of demyelinating diseases. Annu Rev Immunol 10: 153-187. doi: 10.1146/annurev.iy.10.040192.001101
|
| [87] |
Kornek B, Storch MK, Weissert R, et al. (2000) Multiple sclerosis and chronic autoimmune encephalomyelitis: A comparative quantitative study of axonal injury in active, inactive, and remyelinated lesions. Am J Pathol 157: 267-276. doi: 10.1016/S0002-9440(10)64537-3
|
| [88] | Lublin FD, Reingold SC (1996) Defining the clinical course of multiple sclerosis: Results of an international survey. National Multiple Sclerosis Society (USA) Advisory Committee on Clinical Trials of New Agents in Multiple Sclerosis. Neurology 46: 907-911. |
| [89] | Goodkin DE, Reingold S, Sibley W, et al. (1999) Guidelines for clinical trials of new therapeutic agents in multiple sclerosis: Reporting extended results from phase III clinical trials. National Multiple Sclerosis Society Advisory Committee on Clinical Trials of New Agents in Multiple Sclerosis. Ann Neurol 46: 132-134. |
| [90] |
Kappos L, Antel J, Comi G, et al. (2006) Oral fingolimod (FTY720) for relapsing multiple sclerosis. N Eng J Med 355: 1124-1140. doi: 10.1056/NEJMoa052643
|
| [91] | Mehling M, Lindberg R, Raulf F, et al. (2011) Th17 central memory T cells are reduced by FTY720 in patients with multiple sclerosis. Neurology 75: 403-410. |
| [92] |
Kappos L, Radue EW, O’Connor P, et al. (2010) A placebo-controlled trial of oral fingolimod in relapsing multiple sclerosis. N Engl J Med 362: 387-401. doi: 10.1056/NEJMoa0909494
|
| [93] | Saida T, Kikuchi S, Itoyama Y, et al. (2012) A randomized, controlled trial of fingolimod (FTY720) in Japanese patients with multiple sclerosis. Mult Scler 18:1267-1277. |
| [94] |
Kira J, Itoyama Y, Kikuchi S, et al. (2014) Fingolimod (FTY720) therapy in Japanese patients with relapsing multiple sclerosis over 12 months: results of a phase 2 observational extension. BMC Neurology 14: 21. doi: 10.1186/1471-2377-14-21
|
| [95] |
Cohen JA, Barkhof F, Comi G, et al. (2010) Oral fingolimod or intramuscular interferon for relapsing multiple sclerosis. N Engl J Med 362: 402-415. doi: 10.1056/NEJMoa0907839
|
| [96] | Deogracias R, Yazdani M, Dekkers MP, et al (2013) Fingolimod, a sphingosine-1 phosphate receptor modulator, increases BDNF levels and improves symptoms of a mouse model of Rett syndrome. Proc Natl Acad Sci USA 109: 14230-14235. |
| [97] |
Igarashi J, Erwin PA, Dantas AP, et al. (2003) VEGF induces S1P1 receptors in endothelial cells: implications for crosstalk between sphingolipid and growth factors receptors. Proc Natl Acad Sci USA 100: 10664-10669. doi: 10.1073/pnas.1934494100
|
| [98] |
Sanna MG, Liao J, Jo E, et al. (2004). Sphingosine 1-phosphate (S1P) receptor subtypes S1P1 and S1P3, respectively, regulate lymphocyte recirculation and heart rate. J Biol Chem 279: 13839-13848. doi: 10.1074/jbc.M311743200
|
| [99] |
Shimizu H, Takahashi M, T. Kaneko T et al. (2005) KRP-203, a novel synthetic immunosuppressant, prolongs graft survival and attenuates chronic rejection in rat skin and heart allografts. Circulation 111: 222-229. doi: 10.1161/01.CIR.0000152101.41037.AB
|
| [100] |
Hamada M, Nakamura M, Kiuchi M et al. (2010) Removal of sphingosine 1-phosphate receptor-3 (S1P3) agonism is essential, but inadequate to obtain immunomodulating 2-aminopropane-1,3-diol S1P1 agonists with reduced effect on heart rate. J Med Chem 53: 3154-3168. doi: 10.1021/jm901776q
|
| [101] |
Hale JJ, Lynch, CL, Neway WE, et al. (2004) A unique utilization of high throughput screening leads to afford selective, orally bioavailable 1-benzyl-3-carboxyazetidine S1P1 receptor agonists. J Med Chem 47: 6662-6665. doi: 10.1021/jm0492507
|
| [102] |
Vachal P, Toth LM, Hale JJ, et al. (2006) Highly selective and potent agonists of sphingosine-1-phosphate 1 (S1P1) receptor. Bioorg Med Chem Lett 16: 3684-3687. doi: 10.1016/j.bmcl.2006.04.064
|
| [103] | Foss FW, Snyder AH, Davis MD, et al. (2007) Synthesis and biological evaluation of gammma-aminophosphonates as potent, subtype-selective sphingosine 1-phosphate receptor agonists and antagonists. Bioorg Med Chem 15: 663–677. |
| [104] |
Hanessian S, Charron G, Billich A, et al. (2007) Constrained azacyclic analogues of the immunomodulatory agent FTY720 as molecular probes for sphingosine 1-phosphate receptors. Bioorg Med Chem Lett 17: 491-494. doi: 10.1016/j.bmcl.2006.10.014
|
| [105] |
Pan S, Gray NS, Gao W, et al. (2013) Discovery of BAF312 (Siponimod), a Potent and Selective S1P Receptor Modulator. ACS Med Chem Lett 4: 333-337. doi: 10.1021/ml300396r
|
| [106] |
Gergely P, Nuesslein-Hildesheim B, Guerini D, et al. (2012) The selective sphingosine 1-phosphate receptor modulator BAF312 redirects lymphocyte distribution and has species-specific effects on heart rate. Br J Pharmacol 167: 1035-1047. doi: 10.1111/j.1476-5381.2012.02061.x
|
| [107] |
Selmaj K, Li DK, Hartung HP, et al. (2013) Siponimod for patients with relapsing-remitting multiple sclerosis (BOLD): an adaptive, dose-ranging, randomised, phase 2 study. Lancet Neurol 12: 756-767. doi: 10.1016/S1474-4422(13)70102-9
|
| [108] | Biswal S, Veldandi UK, Derne C, et al. (2014) Effect of oral siponimod (BAF312) on the pharmacokinetics and pharmacodynamics of a monophasic oral contraceptive in healthy female subjects. Int J Clin Pharmacol Ther Aug 27. [Epub ahead of print] |
| [109] |
Olsson T, Boster A, Fernández O, et al (2014) Oral ponesimod in relapsing-remitting multiple sclerosis: a randomised phase II trial. J Neurol Neurosurg Psychiatry 85: 1198-1208. doi: 10.1136/jnnp-2013-307282
|
| [110] | Vaclavkova A, Chimenti S, Arenberger P, et al. (2014) Oral ponesimod in patients with chronic plaque psoriasis: a randomised, double-blind, placebo-controlled phase 2 trial. Lancet pii: S0140-6736: 60803-60805. |
| [111] |
Mazurais D, Robert P, Gout B, et al. (2002) Cell type-specific localization of human cardiac S1P receptors. J Histochem Cytochem 50: 661-670. doi: 10.1177/002215540205000507
|
| [112] |
Lukas S, Patnaude L, Haxhinasto S, et al. (2014) No differences observed among multiple clinical S1P1 receptor agonists (functional antagonists) in S1P1 receptor down-regulation and degradation. J Biomol Screen 19: 407-416. doi: 10.1177/1087057113502234
|
| [113] |
Oo ML, Chang SH, Thangada S, et al. (2011) Engagement of S1P1-degradative mechanisms leads to vascular leak in mice. J Clin Invest 121: 2290-2300. doi: 10.1172/JCI45403
|
| [114] |
Violin JD, Crombie AL, Soergel DG, et al. (2014) Biased ligands at G-protein-coupled receptors: promise and progress. Trends Pharmacol Sci 35: 308-316. doi: 10.1016/j.tips.2014.04.007
|
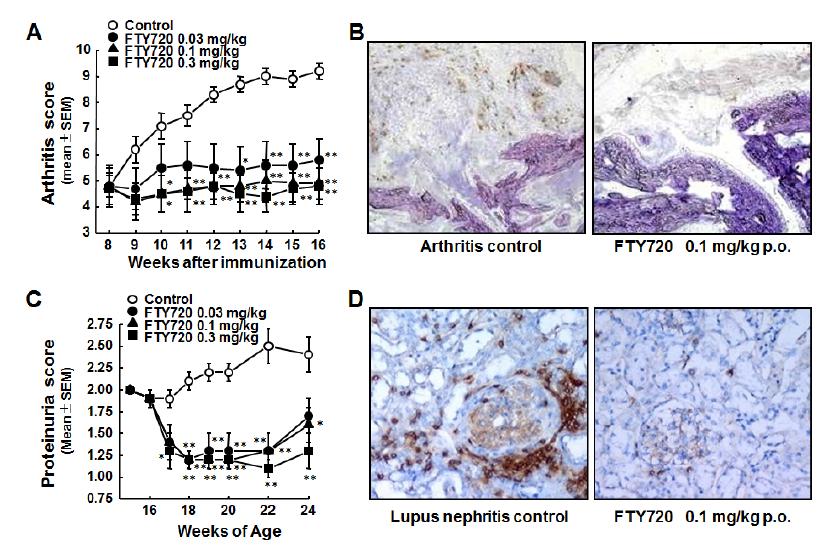
Figures(5)

Kenji Chiba, Yasuhiro Maeda, Noriyasu Seki, Hirotoshi Kataoka, Kunio Sugahara. Role of sphingosine 1-phosphate (S1P) and effects of fingolimod, an S1P receptor 1 functional antagonist in lymphocyte circulation and autoimmune diseases[J]. AIMS Molecular Science, 2014, 1(4): 162-182. doi: 10.3934/molsci.2014.4.162







 DownLoad:
DownLoad: