Citation: J. Amador, D. Armesto, A. Gómez-Corral. Extreme values in SIR epidemic models with two strains and cross-immunity[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 1992-2022. doi: 10.3934/mbe.2019098

| [1] | R. Rowthorn and S. Walther, The optimal treatment of an infectious disease with two strains, J. Math. Biol., 74 (2017), 1753–1791. |
| [2] | L.J.S. Allen, M. Langlais and C.J. Phillips, The dynamics of two viral infections in a single host population with applications to hantavirus, Math. Biosci., 186 (2003), 191–217. |
| [3] | L.J.S. Allen and N. Kirupaharan, Asymptotic dynamics of deterministic and stochastic epidemic models with multiple pathogens, Int. J. Numer. Anal. Mod., 2 (2005), 329–344. |
| [4] | C.P. Bhunu, W. Garira and G. Magombedze, Mathematical analysis of a two strain HIV/AIDS model with antiretroviral treatment, Acta Biotheor., 57 (2009), 361–381. |
| [5] | R. Naresh and A. Tripathi, Modelling and analysis of HIV-TB co-infection in a variable size population, Math. Model. Anal., 10 (2005), 275–286. |
| [6] | M. Nuño, Z. Feng, M. Martcheva, et al., Dynamics of two-strain influenza with isolation and partial cross-immunity, SIAM J. Appl. Math., 65 (2005), 964–982. |
| [7] | M. Lipsitch, C.T. Bergstrom and B.R. Levin, The epidemiology of antibiotic resistance in hospitals: Paradoxes and prescriptions, P. Natl. Acad. Sci., 97 (2000), 1938–1943. |
| [8] | C. Castillo-Chávez and Z. Feng, Mathematical models for the disease dynamics of tuberculosis, in Advances in Mathematical Population Dynamics: Molecular, Cells and Man (eds. O. Avino, D. Axelrod and M. Kimmel), World Scientific, (1998), 629–656. |
| [9] | C. Castillo-Chávez and B. Song, Dynamical models of tuberculosis and their applications, Math. Biosci. Eng., 1 (2004), 361–404. |
| [10] | S. Bhattacharyya, P.H. Gesteland, K. Korgenski, et al., Cross-immunity between strains explains the dynamical pattern of paramyxoviruses, P. Natl. Acad. Sci., 112 (2015), 13396–13400. |
| [11] | W.S. Kendall and I.W. Saunders, Epidemics in competition II: The general epidemic, J. R. Statist. Soc. B, 45 (1983), 238–244. |
| [12] | I.W. Saunders, Epidemics in competition, J. Math. Biol., 11 (1981), 311–318. |
| [13] | A.S. Ackleh and L.J.S. Allen, Competitive exclusion and coexistence for pathogens in an epidemic model with variable population size, J. Math. Biol., 47 (2003), 153–168. |
| [14] | A.S. Ackleh and L.J.S. Allen, Competitive exclusion in SIS and SIR epidemic models with total cross immunity and density-dependent host mortality, Discrete Cont. Dyn.-B, 5 (2005), 175–188. |
| [15] | F. Ball and D. Clancy, The final outcome of an epidemic model with several different types of infective in a large population, J. Appl. Prob., 32 (1995), 579–590. |
| [16] | M.F. Neuts and J.M. Li, An algorithmic study of S-I-R stochastic epidemic models, in Athens Conference on Applied Probability and Time Series Analysis (eds. C.C. Heyde, Y.V. Prohorov, R. Pyke and S.T. Rachev), Lecture Notes in Statistics, Vol. 114, Springer, (1996), 295–306. |
| [17] | D.P. Gaver, P.A. Jacobs and G. Latouche, Finite birth-and-death models in randomly changing environments, Adv. Appl. Probab., 16 (1984), 715–731. |
| [18] | G. Latouche and V. Ramaswami, Introduction to Matrix Analytic Methods in Stochastic Modeling, ASA-SIAM, Philadelphia, 1999. |
| [19] | A. Gómez-Corral and M. López-García, Perturbation analysis in finite LD-QBD processes and applications to epidemic models, Numer. Linear Algebra Appl., (2018);e2160. https://doi.org/10.1002/nla.2160 |
| [20] | J. Amador and M.J. López-Herrero, Cumulative and maximum epidemic sizes for a nonlinear SEIR stochastic model with limited resources, Discrete Contin. Dyn. Syst.-Ser. B, 23 (2018), 2153– 2176. |
| [21] | A. Economou, A. Gómez-Corral and M. López-García, A stochastic SIS epidemic model with heterogeneous contacts, Physica A, 421 (2015), 78–97. |
| [22] | E. Almaraz, A. Gómez-Corral and M.T. Rodríguez-Bernal, On the time to reach a critical number of infections in epidemic models with infective and susceptible immigrants, BioSystems, 144 (2016), 68–77. |
| [23] | E. Almaraz and A. Gómez-Corral, On first-passage times to maximum epidemic sizes in the SIS stochastic model with two different types of infectives, in preparation, (2019). |
| [24] | J.R. Artalejo, A. Gómez-Corral, M. López-García, et al., Stochastic descriptors to study the fate and potential of naive T cell clonotypes in the periphery, J. Math. Biol., 74 (2017), 673–708. |
| [25] | A. Gómez-Corral and M. López-García, Extinction times and size of the surviving species in a two-species competition process, J. Math. Biol., 64 (2012), 255–289. |
| [26] | W.O. Kermack and A.G. McKendrick, A contribution to the mathematical theory of epidemic, Proc. R. Soc. Lon. A, 15 (1927), 700–721. |
| [27] | X. Cen, Z. Feng, Y. Zheng, et al., Bifurcation analysis and global dynamics of a mathematical model of antibiotic resistance in hosptials, J. Math. Biol., 75 (2017), 1463–1485. |
| [28] | T. Vogwill and R.C. MacLean, The genetic basis of the fitness costs of antimicrobial resistance: a meta-analysis approach, Evol. Appl., 8 (2015), 284–295. |
| [29] | T. Britton, Stochastic epidemic models: A survey, Math. Biosci., 225 (2010), 24–35. |
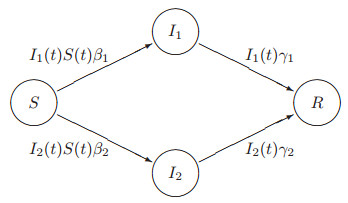
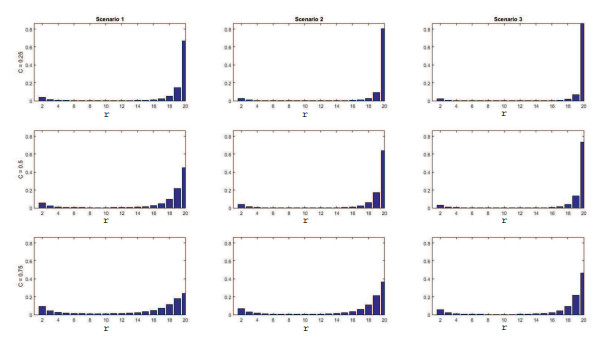
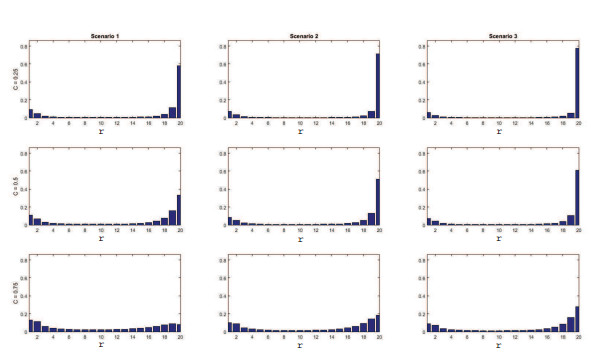
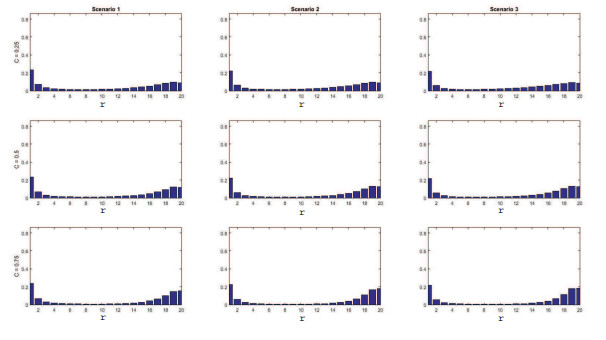
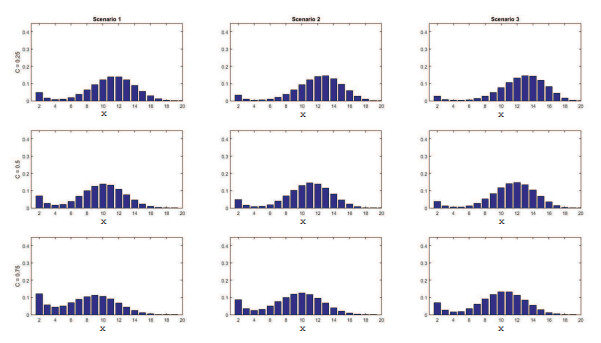
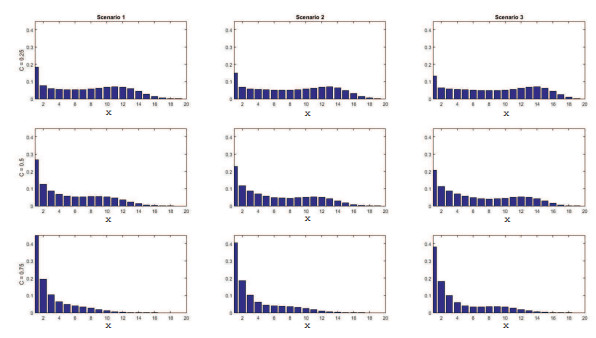
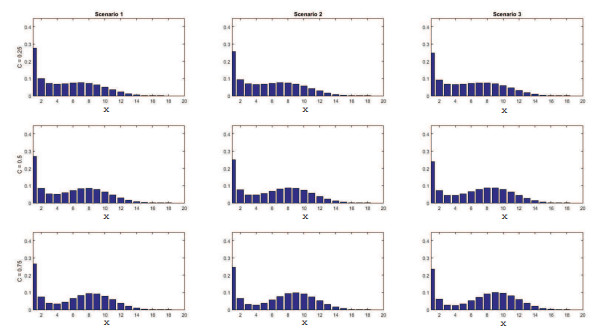
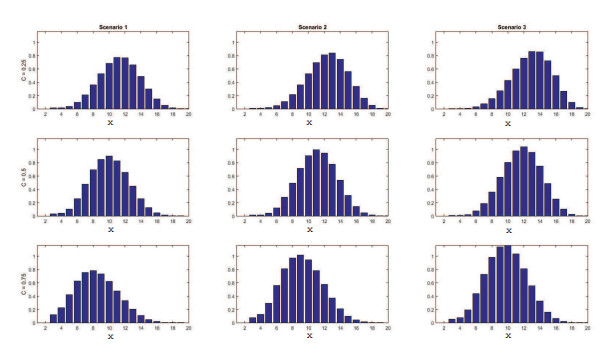
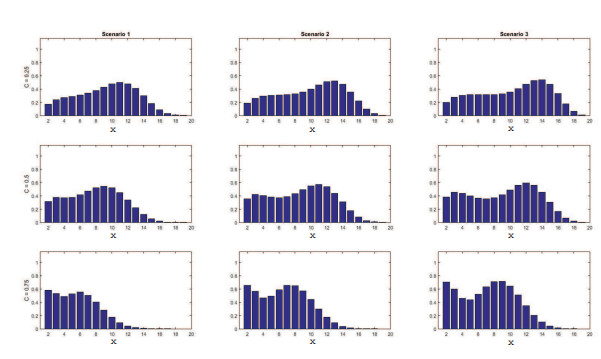
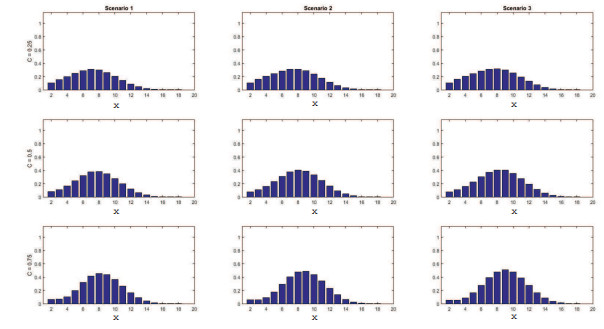
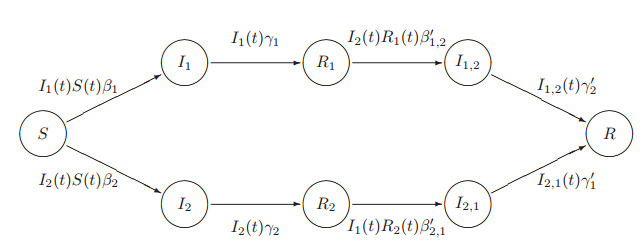
Figures(11) / Tables(4)

J. Amador, D. Armesto, A. Gómez-Corral. Extreme values in SIR epidemic models with two strains and cross-immunity[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 1992-2022. doi: 10.3934/mbe.2019098







 DownLoad:
DownLoad: