Citation: Marcelo F. Pompelli, Gilmara M. Pompelli, Antônio F. M. de Oliveira, Werner C. Antunes. The effect of light and nitrogen availability on the caffeine, theophylline and allantoin contents in the leaves of Coffea arabica L.[J]. AIMS Environmental Science, 2014, 1(1): 1-11. doi: 10.3934/environsci.2013.1.1

| [1] | Hein, L.; Gatzweiler, F. (2006) The economic value of coffee (Coffea arabica) genetic resources. Ecol Econ, 60, 176-185. |
| [2] | DaMatta, F. (2004) Ecophysiological constraints on the production of shaded and unshaded coffee: a review. Field Crops Res, 86, 99-114. |
| [3] | Ashihara, H.; Crozier, A. (2001) Caffeine: a well known but little mentioned compound in plant science. Trends Plant Sci, 6, 407-413. |
| [4] | Waller, G.R.; Ashihara, H.; Kato, M. et al. (2000) Pathways involved in the metabolism of caffeine by Coffea and Camelia plants. In Caffeinated Beverages, Parliment, T.H.; Ho, C.-T.; Schieberle, P., eds, New York: American Chemical Society,. 9-19. |
| [5] | Frischknecht, P.M.; Ulmer-Dufek, J.; Baumann, T.W. (1986) Purine alkaloid formation in buds and developing leaflets of coffea arabica: expression of an optimal defense strategy? Phytochemistry, 25, 613-616. |
| [6] | Zheng, X.Q.; Ashihara, H. (2004) Distribution, biosynthesis and function of purine and pyridine alkaloids in Coffea arabica seedlings. Plant Sci, 166, 807-813. |
| [7] | Suzuki, T.; Waller, G.R. (1984) Biosynthesis and biodegradation of caffeine, theobromine, and theophylline in Coffea arabica L. fruits. J Agric Food Chem, 32, 845-848. |
| [8] | Cordell, G.A. (2013) Fifty years of alkaloid biosynthesis in phytochemistry. Phytochemistry, 91, 29-51. |
| [9] | Ashihara, H.; Sano, H.; Crozier, A. (2008) Caffeine and related purine alkaloids: Biosynthesis, catabolism, function and genetic engineering. Phytochemistry, 69, 841-856. |
| [10] | Vitória, A.P.; Mazzafera, P. (1999) Xanthine degradation and related enzyme activities in leaves and fruits of two coffea species differing in caffeine catabolism. J Agric Food Chem, 47, 1851-1855. |
| [11] | Koyama, Y.; Tomoda, Y.; Kato, M. et al. (2003) Metabolism of purine bases, nucleosides and alkaloids in theobromine-forming Theobroma cacao leaves. Plant Physiol Biochem, 41, 977-984. |
| [12] | Palumbo, M.J.; Putz, F.E.; Talcott, S.T. (2007) Nitrogen fertilizer and gender effects on the secondary metabolism of yaupon, a caffeine-containing North American holly. Oecologia, 151, 1-9. |
| [13] | Gonthier, D.J.; Witter, J.D.; Spongberg, A.L. et al. (2011) Effect of nitrogen fertilization on caffeine production in coffee (Coffea arabica). Chemoecology, 21, 123-130. |
| [14] | Schulthess, B.H.; Baumann, T.W. (1995) Stimulation of caffeine biosynthesis in suspension-cultured coffee cells and the in situ existence of 7-methylxanthosine. Phytochemistry, 38, 1381-1386. |
| [15] | Keller, H.; Wanner, H.; Baumann, T.W. (1972) Caffeine synthesis in fruits and tissue cultures of Coffea arabica. Planta, 108, 339-350. |
| [16] | Cavatte, P.C.; Rodriguez-Lopez, N.F.; Martins, S.C.V. et al. (2012) Functional analysis of the relative growth rate, chemical composition, construction and maintenance costs, and the payback time of Coffea arabica L. leaves in response to light and water availability. J Exp Bot, 63, 3071-3082. |
| [17] | Kim, Y.-S.; Sano, H. (2008) Pathogen resistance of transgenic tobacco plants producing caffeine. Phytochemistry, 69, 882-888. |
| [18] | Bryant, J.P.; Chapin, F.S.; Klein, D.R. (1983) Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory. Oikos, 40, 357-368. |
| [19] | Pompelli, M.F.; Pompelli, G.M.; Cabrini, E.C. et al. (2012) Leaf anatomy, ultrastructure and plasticity of Coffea arabica L. in response to light and nitrogen availability. Biotemas, 25, 13-28. |
| [20] | Hamilton, J.G.; Zangerl, A.R.; DeLucia, E.H. et al. (2001) The carbon-nutrient balance hypothesis: its raise and fall. Ecol Lett, 4, 86-95. |
| [21] | Mazzafera, P. (1999) Mineral nutrition and caffeine content in coffee leaves. Bragantia, 58, 387-391. |
| [22] | Mazzafera, P.; Gonçalves, K.V. (1999) Nitrogen compounds in the xylem sap of coffee. Phytochemistry, 50, 383-386. |
| [23] | Pompelli, M.F.; Martins, S.C.V.; Antunes, W.C. et al. (2010) Photosynthesis and photoprotection in coffee leaves is affected by nitrogen and light availabilities in winter conditions. J Plant Physiol, 167, 1052-1060. |
| [24] | Epstein, E. (1972) Mineral nutrition of plants: principles and perspectives, New York: John Wiley & Sons. |
| [25] | Moraes, F.R.P. (1981) Adubação do cafeeiro: macronutrientes e adubação orgânica. In Nutrição e adubação do cafeeiro, Malavolta, E.; Yamada, T.; Guidolin, J.A., eds, Piracicaba: Instituto Internacional da Potassa, 77-89. |
| [26] | Filippi, S.B.; Azevedo, R.A.; Sodek, L. et al. (2007) Allantoin has a limited role as nitrogen source in cultured coffee cells. J Plant Physiol, 164, 544-552. |
| [27] | Praxedes, S.C.; DaMatta, F.M.; Loureiro, M.E. et al. (2006) Effects of long-term soil drought on photosynthesis and carbohydrate metabolism in mature robusta coffee (Coffea canephora Pierre var. kouillou) leaves. Environ Exp Bot, 56, 263-273. |
| [28] | Bradford, M. (1976) Rapid and quantitative method for quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anual Biochem, 72, 284-252. |
| [29] | Pompelli, M.F.; Barata-Luís, R.M.; Vitorino, H.S. et al. (2010) Photosynthesis, photoprotection and antioxidant activity of purging nut under drought deficit and recovery. Biomass Bioenerg, 34, 1207-1215. |
| [30] | Waller, G.R.; Jurzysta, M.; Karns, T.K.B. et al. (1991) Isolation and identification of ursolic acid from Coffea arabica L. (Coffee) leaves. Asic, 14, 245-257. |
| [31] | Coelho, G.C.; Rachwal, M.F.G.; Dedecek, R.A. et al. (2007) Effect of light intensity on methylxanthine contents of Ilex paraguariensis A. St. Hil. Biochem Syst Ecol, 35, 75-80. |
| [32] | Song, R.; Kelman, D.; Johns, K.L. et al. (2012) Correlation between leaf age, shade levels, and characteristics beneficial natural constituints of tea (Camelia sinensis) grown in Hawaii. Food Chem, 133, 707-714. |
| [33] | Koshiishi, C.; Ito, E.; Kato, A. et al. (2000) Purine alkaloid biosynthesis in young leaves of Camellia sinensis in light and darkness. J Plant Res, 113, 217-221. |
| [34] | Kato, A.; Crozier, A.; Ashihara, H. (1998) Subcellular localization of the N-3 methyltransferase involved in caffeine biosynthesis in tea. Phytochemistry, 48, 777-779. |
| [35] | Kato, M.; Mizuno, K.; Fujimura, T. et al. (1999) Purification and characterization of caffeine synthase from tea leaves. Plant Physiol, 120, 579-586. |
| [36] | Allen, J.F. (2005) A redox swith hypothesis for the origin of two light reactions in photosynthesis. FEBS Lett, 579, 963-968. |
| [37] | Stitt, M.; Krapp, A. (1999) The interaction between elevated carbon dioxide and nitrogen nutrition: the physiological and molecular background. Plant Cell Environ, 22, 583-621. |
| [38] | Chapin, F.S.; Schulze, E.; Mooney, H.A. (1990) The ecology and economics of storage in plants. Annu Rev Ecol Syst, 21, 423-447. |
| [39] | Pugnaire, F.I.; Valladares, F. (2007) Functional plant ecology 2nd ed, CRCpress, New York, |
| [40] | Campbell, W.H. (1999) Nitrate reductase structure, function and regulation: bridging the gap between biochemistry and physiology. Annu Rev Plant Phys, 50, 277-303. |
| [41] | Meguro, N.E.; Magalhães, A.C. (1982) Atividade da redutase de nitrato em cultivares de café. Pesqui Agropecu Bras, 17, 1725-1731. |
| [42] | Carelli, M.L.C.; Fahl, J.I.; Magalhães, A.C. (1990) Atividade da redutase de nitrato em folhas e raízes de plantas de café (Coffea arabica L.). Rev Bras Bot, 13, 119-123. |
| [43] | Oliveira, I.; Brears, T.; Knight, T. et al. (2002) Overexpression of cytosolic glutamine synthetase relation to nitrogen, light, and photorespiration. Plant Physiol, 129, 1170-1180. |
| [44] | Waller, G.R.; Anaya-Lang, A.L.; Sagrero-Nieves, L. et al. (1989) A problem in coffee plantations: autotoxicity of caffeine and other compounds. Asic, 13, 363-371. |
| [45] | Yamaoka-Yano, D.M.; Mazzafera, P. (1997) Degradation of caffeine by Pseudomonas putida isolated from soil. Allelopathy J, 5, 23-34. |
| [46] | Beltrán, J.G.; Leask, R.L.; Brown, W.A. (2006) Activity and stability of caffeine demethylases found in Pseudomonas putida IF-3. Biochem Eng J., 31, 8-13. |
| [47] | Koshiro, Y.; Zheng, X.-Q.; Wang, M.-L. et al. (2006) Changes in content and biosynthetic activity of caffeine and trigonelline during growth and ripening of Coffea arabica and Coffea canephora fruits. Plant Sci, 171, 242-250. |
| [48] | Nazario, G.M.; Lovatt, C.J. (1993) Regulation of purine metabolism in intact leaves of Coffea arabica. Plant Physiol, 103, 1195-1201. |
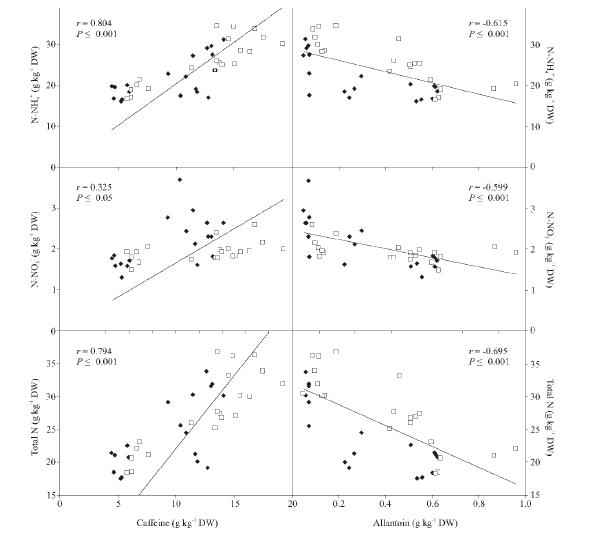
Figures(1) / Tables(1)

Marcelo F. Pompelli, Gilmara M. Pompelli, Antônio F. M. de Oliveira, Werner C. Antunes. The effect of light and nitrogen availability on the caffeine, theophylline and allantoin contents in the leaves of Coffea arabica L.[J]. AIMS Environmental Science, 2014, 1(1): 1-11. doi: 10.3934/environsci.2013.1.1







 DownLoad:
DownLoad: