Citation: Ernest Greene. New encoding concepts for shape recognition are needed[J]. AIMS Neuroscience, 2018, 5(3): 162-178. doi: 10.3934/Neuroscience.2018.3.162

| [1] | Boorstein DJ (1983) The Discoverers, New York: Random House. |
| [2] |
Hubel DH, Wiesel TN (1959) Receptive fields of single neurons in the cat's striate cortex. J Physiol 148: 574–591. doi: 10.1113/jphysiol.1959.sp006308

|
| [3] |
Hubel DH, Wiesel TN (1968) Receptive fields and functional architecture of monkey striate cortex. J Physiol 195: 215–243. doi: 10.1113/jphysiol.1968.sp008455
|
| [4] |
Sceniak MP, Hawken MJ, Shapley R (2001) Visual spatial characterization of macaque V1 neurons. J Neurophysiol 85: 1873–1887. doi: 10.1152/jn.2001.85.5.1873
|
| [5] |
Greene E (2007) Recognition of objects displayed with incomplete sets of discrete boundary dots. Perceptual Mot Skills 104: 1043–1059. doi: 10.2466/pms.104.4.1043-1059
|
| [6] |
Greene E (2007) The integration window for shape cues as a function of ambient illumination. Behav Brain Funct 3: 15. doi: 10.1186/1744-9081-3-15
|
| [7] |
Greene E (2015) Evaluating letter recognition, flicker fusion, and the talbot-plateau law using microsecond-duration flashes. PLoS One 10: e0123458. doi: 10.1371/journal.pone.0123458
|
| [8] |
Greene E (2016) Recognizing words and reading sentences with microsecond flash displays. PLoS One 11: e0145697. doi: 10.1371/journal.pone.0145697
|
| [9] |
Greene E (2008) Additional evidence that contour attributes are not essential cues for object recognition. Behav Brain Funct 4: e26. doi: 10.1186/1744-9081-4-26
|
| [10] | Greene E (2016) How do we know whether three dots form an equilateral triangle? JSM Brain Sci 1: 1002. |
| [11] |
Fukushima K (1980) Neocognitron: A self-organizing neural network model for a mechanism of pattern recognition unaffected by shift in position. Biol Cybern 36: 193–202. doi: 10.1007/BF00344251
|
| [12] |
Rolls ET (1992) Neurophysiological mechanisms underlying face processing within and beyond the temporal cortical visual areas. Phil Trans R Soc 335: 11–21. doi: 10.1098/rstb.1992.0002
|
| [13] |
Wallis G, Rolls ET (1997) Invariant face and object recognition in the visual system. Prog Neurobiol 51: 167–194. doi: 10.1016/S0301-0082(96)00054-8
|
| [14] |
Rodríguezsánchez AJ, Tsotsos JK (2012) The roles of endstopped and curvature tuned computations in a hierarchical representation of 2D shape. PLoS One 7: e42058. doi: 10.1371/journal.pone.0042058
|
| [15] |
Riesenhuber M, Poggio T (2000) Models of object recognition. Nature Neurosci Suppl 3: 1199–1204. doi: 10.1038/81479
|
| [16] |
Pasupathy A, Connor CE (2001) Shape representation in area V4: Position-specific tuning for boundary conformation. J Neurophysiol 86: 2505–2519. doi: 10.1152/jn.2001.86.5.2505
|
| [17] |
Suzuki N, Hashimoto N, Kashimori Y, et al. (2004) A neural model of predictive recognition in form pathway of visual cortex. Bio Systems 76: 33–42. doi: 10.1016/j.biosystems.2004.05.004
|
| [18] |
Pinto N, Cox DD, DeCarlo JJ (2008) Why is real-world visual object recognition hard? PLoS Comput Biol 4: e27. doi: 10.1371/journal.pcbi.0040027
|
| [19] |
Greene E, Hautus MJ (2017) Demonstrating invariant encoding of shapes using a matching judgment protocol. AIMS Neurosci 4: 120–147. doi: 10.3934/Neuroscience.2017.3.120
|
| [20] |
Greene E, Hautus MJ (2018) Evaluating persistence of shape information using a matching protocol. AIMS Neurosci 5:81-96. doi: 10.3934/Neuroscience.2018.1.81
|
| [21] | Greene E, Onwuzulike O (2017) What constitutes elemental shape information for biological vision? Trends Artif Intell 1: 22–26. |
| [22] | Greene E (2018) Rapid de novo shape encoding: A challenge to connectionist modeling. arXiv: 1801.02256v1 |
| [23] | Karplus I, Goren M, Algorn D (1982) A preliminary experimental analysis of predator face recognition by Chromis caenuleus (Pisces, Pomacentridae). Z Tierpsychol 58: 53–65. |
| [24] |
Siebeck UE, Parker AN, Sprenger D, et al. (2010) A species of reef fish that uses untraviolet pattterns for covert face recognition. Curr Biol 20: 407–410. doi: 10.1016/j.cub.2009.12.047
|
| [25] |
Karplus I (2006) Predator recognition and social facilitation of predator avoidance in coral reef fish Dascyllus marginatus juveniles. Mar Ecol Prog Ser 319: 215–223. doi: 10.3354/meps319215
|
| [26] |
Siebeck UE, Litherland L, Wallis GM (2009) Shape learning and discrimination in reef fish. J Exp Biol 212: 2113–2119. doi: 10.1242/jeb.028936
|
| [27] |
Newport C, Wallis G, Reshitnyk Y, et al. (2016) Discrimination of human faces by archerfish (Toxotes catareus). Sci Rep 6: 27523. doi: 10.1038/srep27523
|
| [28] |
Abbas F, Martin MP (2014) Fish vision: Size selectivity in the zebrafish retinotectal pathway. Curr Biol 24: 1048–1050. doi: 10.1016/j.cub.2014.09.043
|
| [29] |
Greene E, Waksman P (1987) Grid analysis: Continuing the search for a metric of shape. J Math Psychol 31: 338–365. doi: 10.1016/0022-2496(87)90020-4
|
| [30] | Waksman P, Greene E (1988) Optical image encoding and comparing using scan autocorrelation. United States Patent 4745633. |
| [31] | Greene E (2010) Encoding system providing discrimination, classification, and recognition of shapes and patterns. United States Patent 7809195. |
| [32] | Greene E (2016) Retinal encoding of shape boundaries. JSM Anat Physiol 1: e1002. |
| [33] |
Dacey DM (1989) Axon-bearing amacrine cells of the Macaque monkey retina. J Comp Neurol 284: 275–293. doi: 10.1002/cne.902840210
|
| [34] | Rodieck RW, (1998) The primate retina, In: Steklis HD, Erwin J (eds), Comparative Primate Biology, Vol. 4, Neuroscience, New York, 203–278. |
| [35] | Ammermuller J, Weller R (1988) Physiological and morphological characterization of OFF-center amacrine cells in the turtle retina. J Comp Physiol 273: 137–148. |
| [36] |
Famiglietti EV (1992) Polyaxonal amacrine cells of rabbit retina: Morphology and stratification of PA1 cells. J Comp Neurol 316: 391–405. doi: 10.1002/cne.903160402
|
| [37] |
Famiglietti EV (1992) Polyaxonal amacrine cells of rabbit retina: Size and distribution of PA1 cells. J Comp Neurol 316: 406–421. doi: 10.1002/cne.903160403
|
| [38] |
Freed MA, Pflug R, Kolb H, et al. (1996) ON-OFF amacrine cells in cate retina. J Comp Neural 364: 556–566. doi: 10.1002/(SICI)1096-9861(19960115)364:3<556::AID-CNE12>3.0.CO;2-N
|
| [39] |
Volgi B, Xin D, Amarillo Y, et al. (2001) Morphology and physiology of the polyaxonal amacrine cells in the rabbit retina. J Comp Neurol 440: 109–125. doi: 10.1002/cne.1373
|
| [40] |
Wright LL, Vaney DI (2004) The type 1 polyaxonal amacrine cells of the rabbit retina: A tracer-coupled study. Visual Neurosci 21: 145–155. doi: 10.1017/S0952523804042063
|
| [41] |
Greene E (2007) Retinal encoding of ultrabrief shape recognition cues. PLoS One 2: e871. doi: 10.1371/journal.pone.0000871
|
| [42] |
Nordberg H, Hautus M, Greene E (2018) Visual encoding of partial unknown shape boundaries. AIMS Neurosci 5: 132–147. doi: 10.3934/Neuroscience.2018.2.132
|
| [43] |
Bullock TH (1993) Integrative systems research in the brain: Resurgence and new opportunities. Ann Rev Neurosci 16: 1–15. doi: 10.1146/annurev.ne.16.030193.000245
|
| [44] |
Hopfield JJ (1995) Pattern recognition computation using action potential timing for stimulus representation. Nature 376: 33–36. doi: 10.1038/376033a0
|
| [45] |
Thorpe S, Fize D, Marlot C (1996) Speed of processing in the human visual system. Nature 381: 520–522. doi: 10.1038/381520a0
|
| [46] |
Thorpe S, Delorme A, VanRullen R (2001) Spike-based strategies for rapid processing. Neural Net 14: 715–725. doi: 10.1016/S0893-6080(01)00083-1
|
| [47] | Vanrullen R, Thorpe SJ (2001) Is it a bird? Is it a plane? Ultra-rapid visual categorisation of natural and artifactual objects. Perception 30: 655–688. |
| [48] |
Vanrullen R, Thorpe SJ (2002) Surfing a spike wave down the ventral stream. Vision Res 42: 2593–2615. doi: 10.1016/S0042-6989(02)00298-5
|
| [49] | Ahissar E, Arieli A (2012) Seeing via miniature eye movements: A dynamic hypothesis for vision. Front Comput Neurosci 6: e89. |
| [50] |
Rucci M, Victor JD (2015) The unsteady eye: An information-processing stage, not a bug. Trends Neurosci 38: 195–205. doi: 10.1016/j.tins.2015.01.005
|
| [51] |
Gollisch T, Meister M (2008) Rapid neural coding in the retina with relative spike latencies. Science 319: 1108–1111. doi: 10.1126/science.1149639
|
| [52] | Greene E, Patel Y (2018) Scan encoding of two-dimensional shapes as an alternative neuromorphic concept. Trends Artific Intell 1: 27–33. |
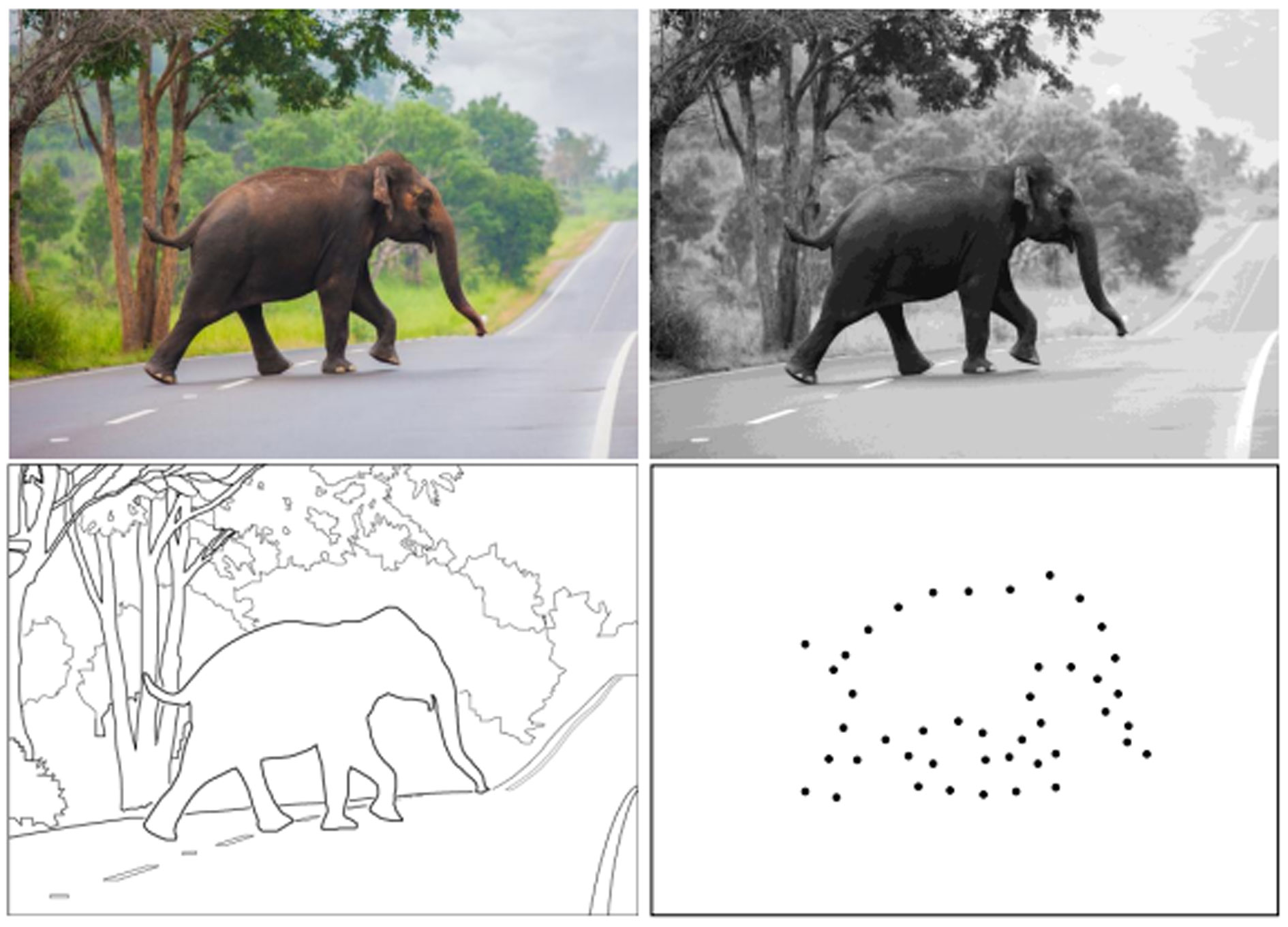
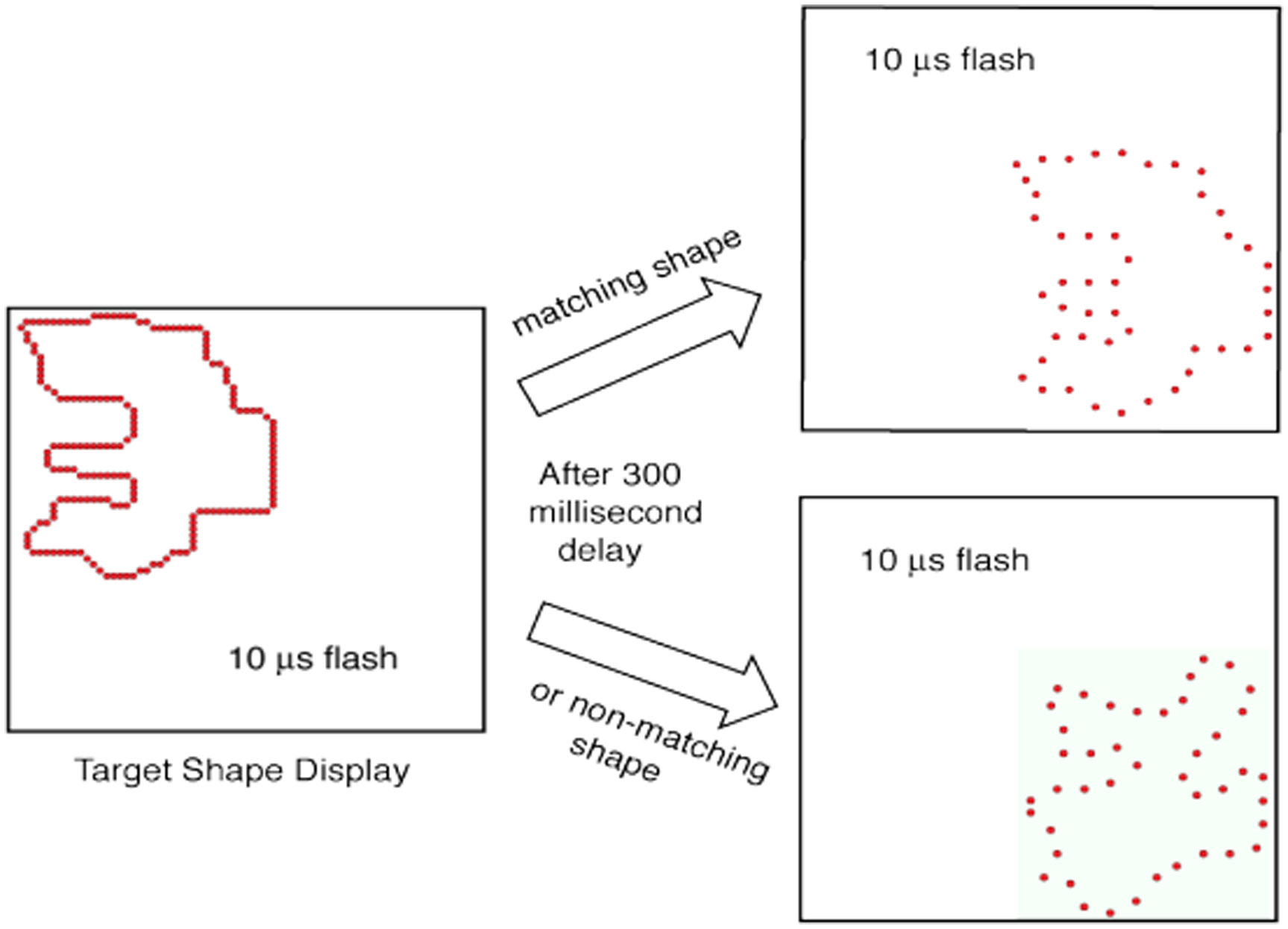
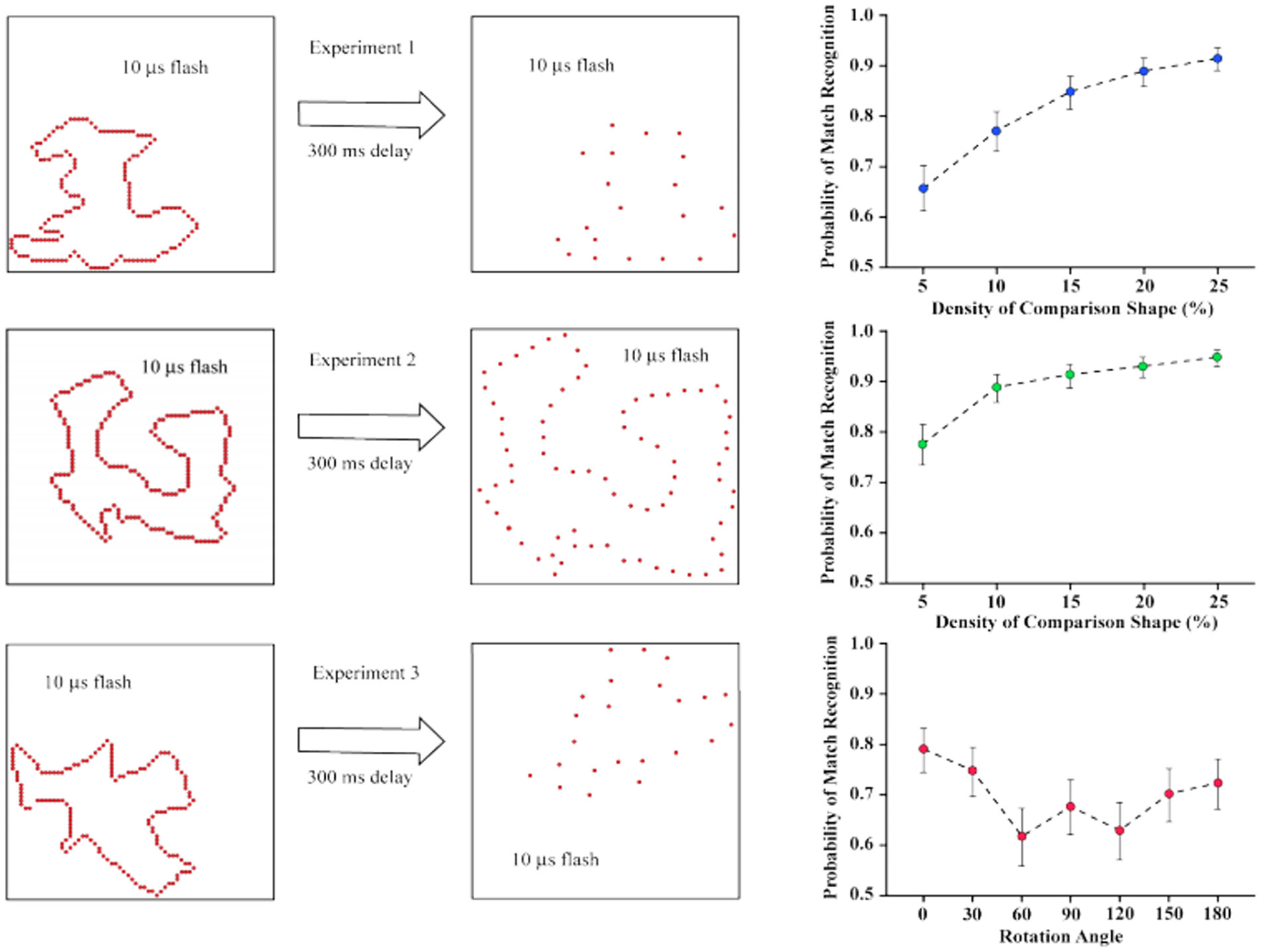
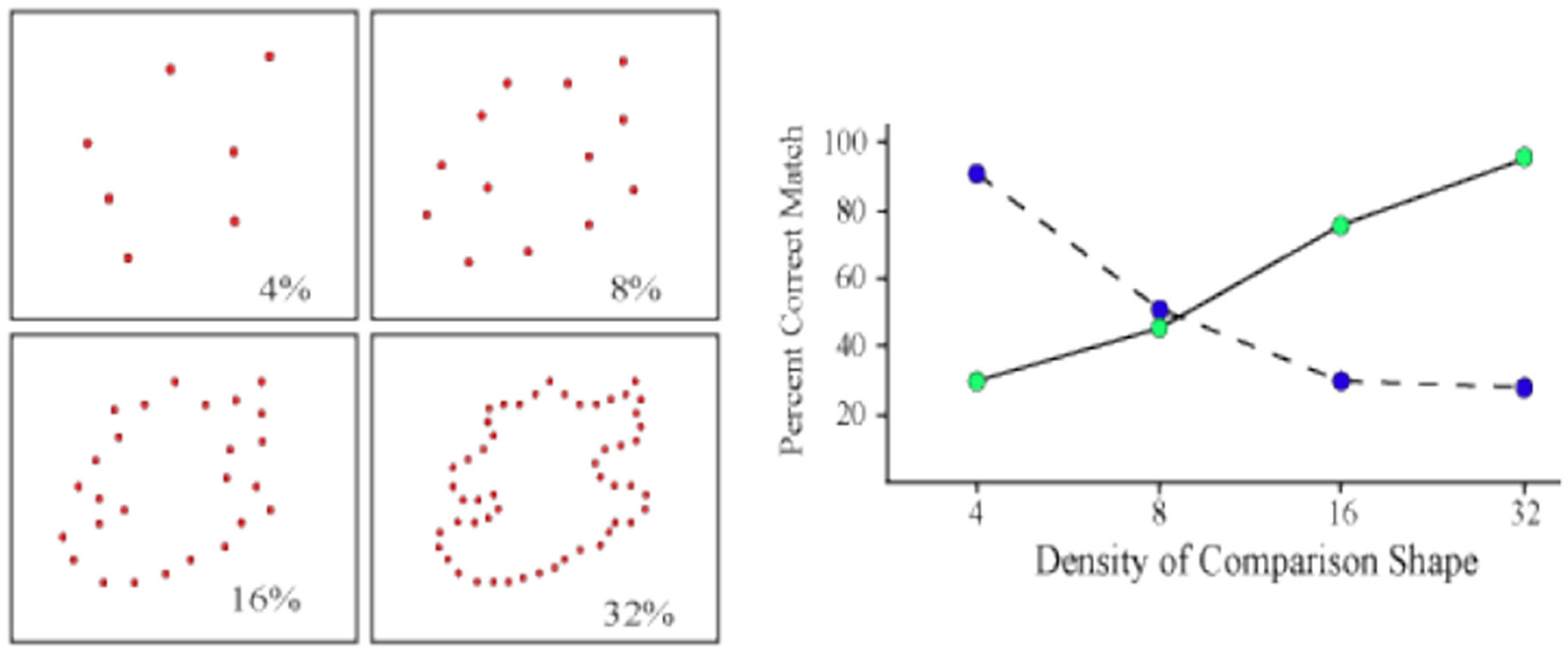

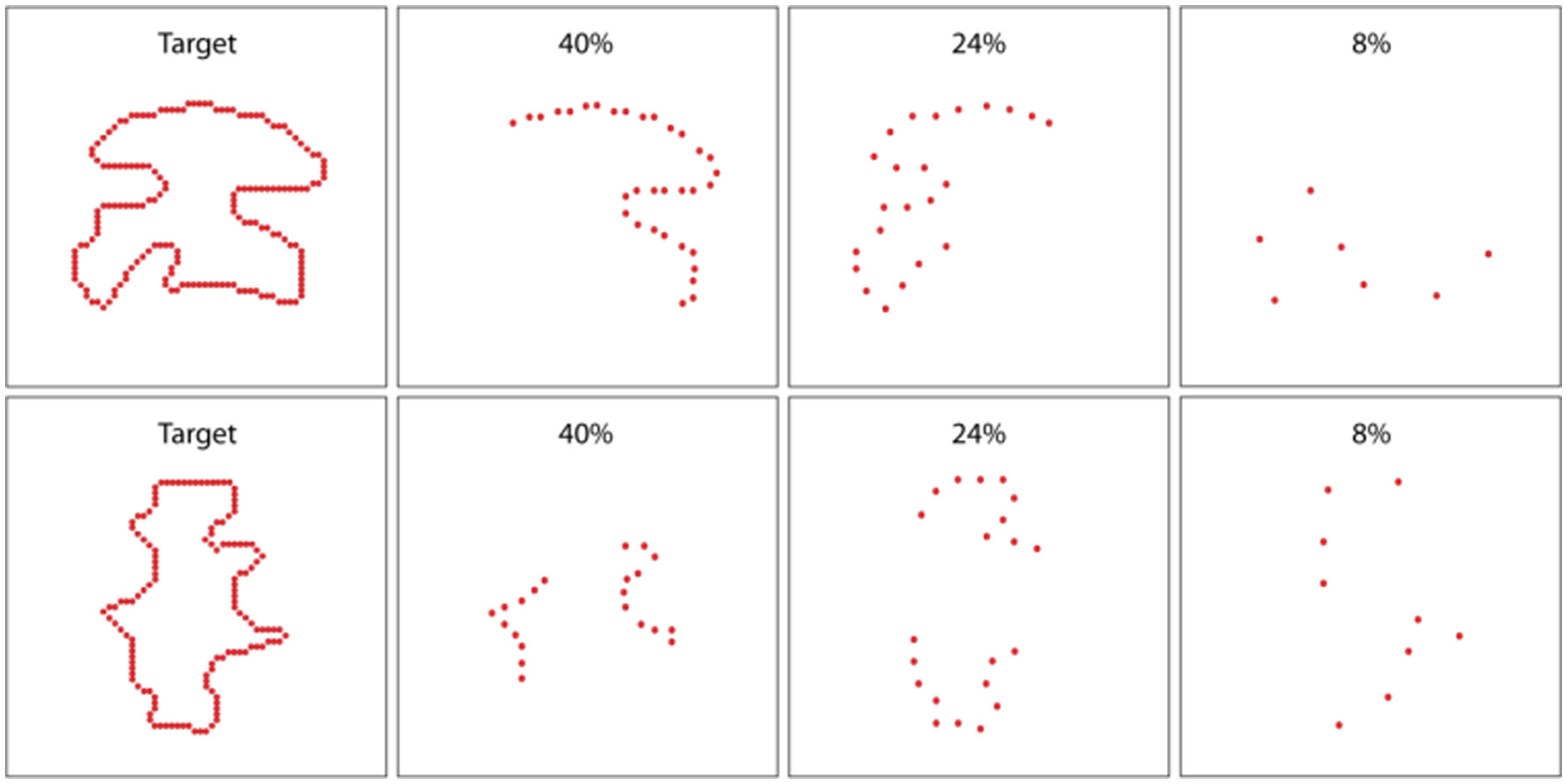
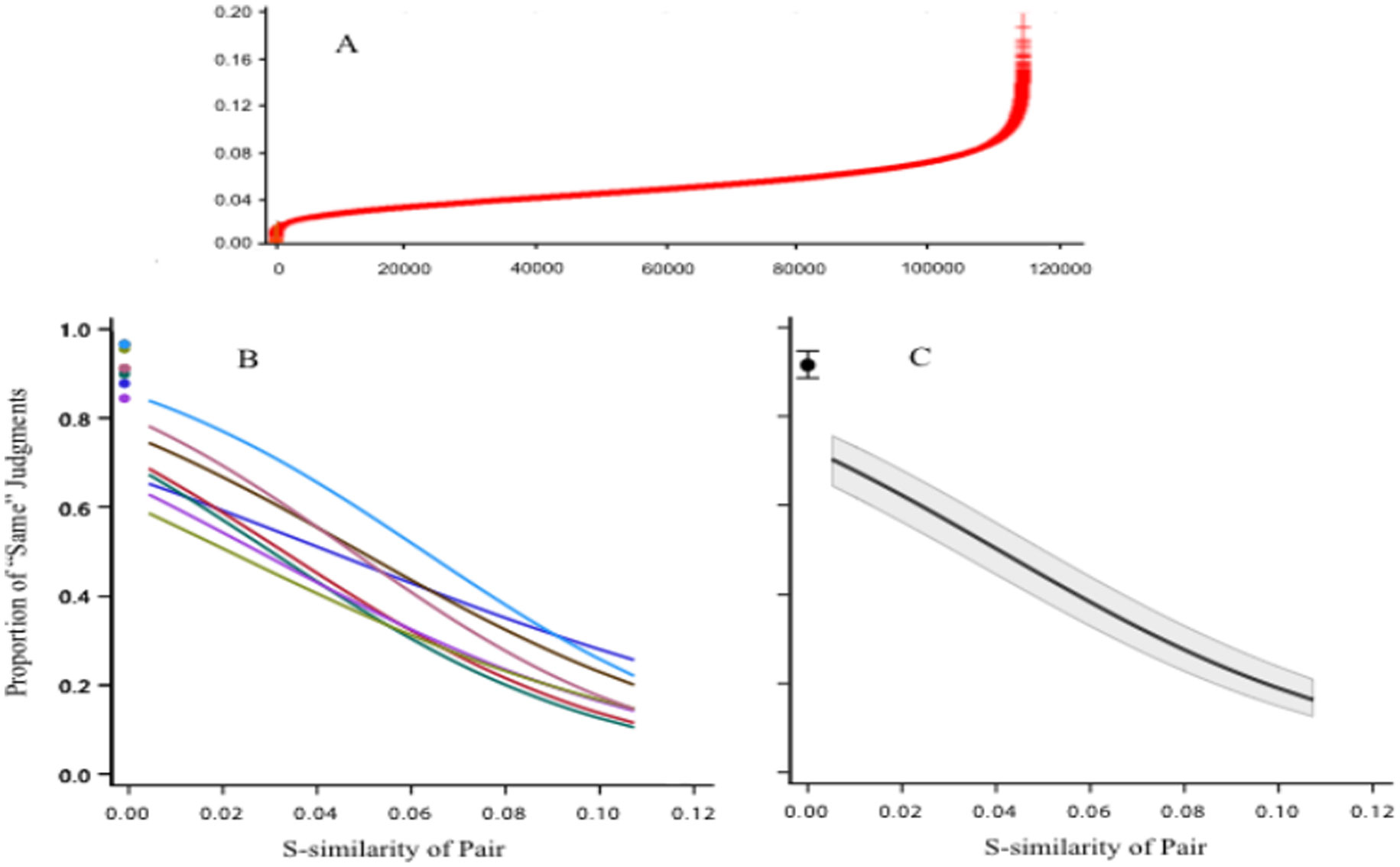
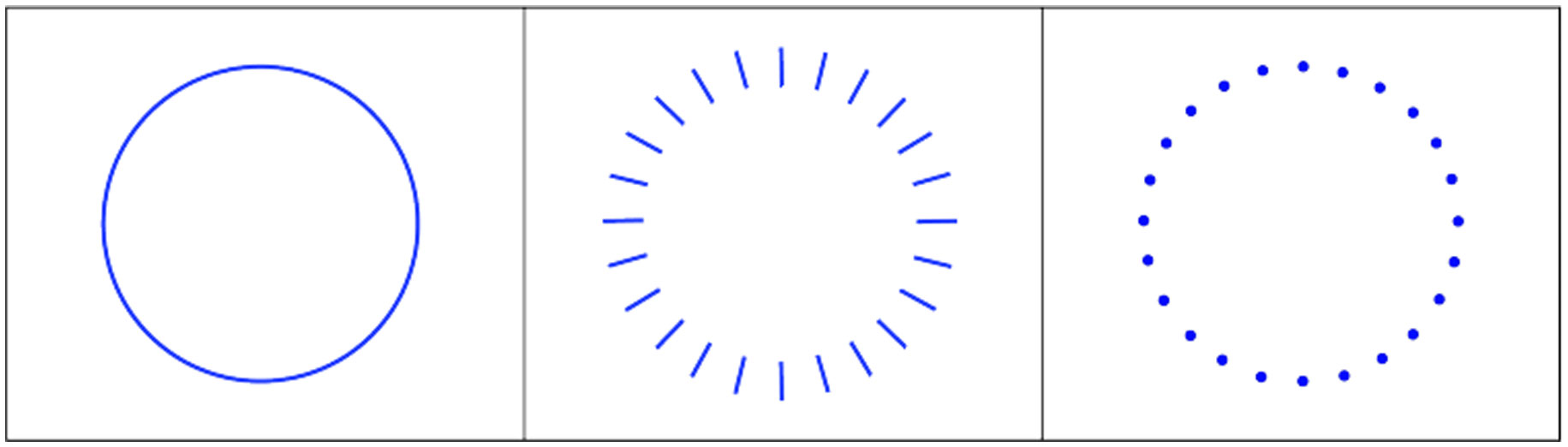
Figures(9)

Ernest Greene. New encoding concepts for shape recognition are needed[J]. AIMS Neuroscience, 2018, 5(3): 162-178. doi: 10.3934/Neuroscience.2018.3.162







 DownLoad:
DownLoad: