Citation: Omer Qibi, Severine Audusseau, Andrea Mogas, Zoulfia Allakhverdi, Abdelilah Soussi Gounni, Saba Al Heialy, Qutayba Hamid. No evidence for IgE receptor FcεRI expression on bronchial epithelial cells of asthmatic patients[J]. AIMS Allergy and Immunology, 2018, 2(4): 165-179. doi: 10.3934/Allergy.2018.4.165

| [1] |
Galli SJ, Tsai M, Piliponsky AM (2008) The development of allergic inflammation. Nature 454: 445–454. doi: 10.1038/nature07204

|
| [2] |
James AL, Wenzel S (2007) Clinical relevance of airway remodelling in airway diseases. Eur Respir J 30: 134–155. doi: 10.1183/09031936.00146905
|
| [3] |
Sumi Y, Hamid Q (2007) Airway remodeling in asthma. Allergol Int 56: 341–348. doi: 10.2332/allergolint.R-07-153
|
| [4] |
Crimi E, Scordamaglia A, Crimi P, et al. (1983) Total and specific IgE in serum, bronchial lavage and bronchoalveolar lavage of asthmatic patients. Allergy 38: 553–559. doi: 10.1111/j.1398-9995.1983.tb04139.x
|
| [5] |
Platts-Mills TA (2001) The role of immunoglobulin E in allergy and asthma. Am J Respir Crit Care Med 164: S1–S5. doi: 10.1164/ajrccm.164.supplement_1.2103024
|
| [6] |
Gould HJ, Sutton BJ (2008) IgE in allergy and asthma today. Nat Rev Immunol 8: 205–217. doi: 10.1038/nri2273
|
| [7] |
Holgate S, Casale T, Wenzel S, et al. (2005) The anti-inflammatory effects of omalizumab confirm the central role of IgE in allergic inflammation. J Allergy Clin Immunol 115: 459–465. doi: 10.1016/j.jaci.2004.11.053
|
| [8] |
Humbert M, Busse W, Hanania NA, et al. (2014) Omalizumab in asthma: An update on recent developments. J Allergy Clin Immunol Pract 2: 525–536. doi: 10.1016/j.jaip.2014.03.010
|
| [9] |
Holowka D, Sil D, Torigoe C, et al. (2007) Insights into immunoglobulin E receptor signaling from structurally defined ligands. Immunol Rev 217: 269–279. doi: 10.1111/j.1600-065X.2007.00517.x
|
| [10] |
Kraft S, Kinet JP (2007) New developments in FcepsilonRI regulation, function and inhibition. Nat Rev Immunol 7: 365–378. doi: 10.1038/nri2072
|
| [11] |
Galli SJ, Tsai M (2012) IgE and mast cells in allergic disease. Nat Med 18: 693–704. doi: 10.1038/nm.2755
|
| [12] | Maurer D, Fiebiger S, Ebner C, et al. (1996) Peripheral blood dendritic cells express Fc epsilon RI as a complex composed of Fc epsilon RI alpha- and Fc epsilon RI gamma-chains and can use this receptor for IgE-mediated allergen presentation. J Immunol 157: 607–616. |
| [13] |
Redhu NS, Gounni AS (2013) The high affinity IgE receptor (FcepsilonRI) expression and function in airway smooth muscle. Pulm Pharmacol Ther 26: 86–94. doi: 10.1016/j.pupt.2012.04.004
|
| [14] |
Untersmayr E, Bises G, Starkl P, et al. (2010) The high affinity IgE receptor Fc epsilonRI is expressed by human intestinal epithelial cells. PLoS One 5: e9023. doi: 10.1371/journal.pone.0009023
|
| [15] |
Yokota A, Kikutani H, Tanaka T, et al. (1988) Two species of human Fc epsilon receptor II (Fc epsilon RII/CD23): Tissue-specific and IL-4-specific regulation of gene expression. Cell 55: 611–618. doi: 10.1016/0092-8674(88)90219-X
|
| [16] |
Foster B, Metcalfe DD, Prussin C (2003) Human dendritic cell 1 and dendritic cell 2 subsets express FcepsilonRI: Correlation with serum IgE and allergic asthma. J Allergy Clin Immunol 112: 1132–1138. doi: 10.1016/j.jaci.2003.09.011
|
| [17] |
Sihra BS, Kon OM, Grant JA, et al. (1997) Expression of high-affinity IgE receptors (Fc epsilon RI) on peripheral blood basophils, monocytes, and eosinophils in atopic and nonatopic subjects: Relationship to total serum IgE concentrations. J Allergy Clin Immunol 99: 699–706. doi: 10.1016/S0091-6749(97)70033-2
|
| [18] | Malveaux FJ, Conroy MC, Jr AN, et al. (1978) IgE receptors on human basophils. Relationship to serum IgE concentration. J Clin Invest 62: 176–181. |
| [19] |
Nopp A, Johansson SG, Adedoyin J, et al. (2010) After 6 years with Xolair; a 3-year withdrawal follow-up. Allergy 65: 56–60. doi: 10.1111/j.1398-9995.2009.02144.x
|
| [20] |
Molimard M, Mala L, Bourdeix I, et al. (2014) Observational study in severe asthmatic patients after discontinuation of omalizumab for good asthma control. Respir Med 108: 571–576. doi: 10.1016/j.rmed.2014.02.003
|
| [21] |
Roth M, Zhong J, Zumkeller C, et al. (2013) The role of IgE-receptors in IgE-dependent airway smooth muscle cell remodelling. PLoS One 8: e56015. doi: 10.1371/journal.pone.0056015
|
| [22] |
Campbell AM, Vachier I, Chanez P, et al. (1998) Expression of the high-affinity receptor for IgE on bronchial epithelial cells of asthmatics. Am J Respir Cell Mol Biol 19: 92–97. doi: 10.1165/ajrcmb.19.1.2648
|
| [23] |
Takhar P, Corrigan CJ, Smurthwaite L, et al. (2007) Class switch recombination to IgE in the bronchial mucosa of atopic and nonatopic patients with asthma. J Allergy Clin Immunol 119: 213–218. doi: 10.1016/j.jaci.2006.09.045
|
| [24] |
Bajpai R, Lesperance J, Kim M, et al. (2008) Efficient propagation of single cells accutase-dissociated human embryonic stem cells. Mol Reprod Dev 75: 818–827. doi: 10.1002/mrd.20809
|
| [25] |
Furuichi K, Rivera J, Isersky C (1985) The receptor for immunoglobulin E on rat basophilic leukemia cells: Effect of ligand binding on receptor expression. Proc Natl Acad Sci USA 82: 1522–1525. doi: 10.1073/pnas.82.5.1522
|
| [26] |
Gebhardt T, Lorentz A, Detmer F, et al. (2005) Growth, phenotype, and function of human intestinal mast cells are tightly regulated by transforming growth factor beta1. Gut 54: 928–934. doi: 10.1136/gut.2004.054650
|
| [27] |
Gomez G, Ramirez CD, Rivera J, et al. (2005) TGF-beta 1 inhibits mast cell Fc epsilon RI expression. J Immunol 174: 5987–5993. doi: 10.4049/jimmunol.174.10.5987
|
| [28] |
Okayama Y, Okumura S, Sagara H, et al. (2009) FcepsilonRI-mediated thymic stromal lymphopoietin production by interleukin-4-primed human mast cells. Eur Respir J 34: 425–435. doi: 10.1183/09031936.00121008
|
| [29] |
Corren J, Kavati A, Ortiz B, et al. (2018) Patient-reported outcomes in moderate-to-severe allergic asthmatics treated with omalizumab: A systematic literature review of randomized controlled trials. Curr Med Res Opin 34: 65–80. doi: 10.1080/03007995.2017.1395734
|
| [30] | Jr MGD, Bochner BS, Adelman DC, et al. (1997) Down-regulation of Fc(epsilon)RI expression on human basophils during in vivo treatment of atopic patients with anti-IgE antibody. J Immunol 158: 1438–1445. |
| [31] |
Djukanovic R, Wilson SJ, Kraft M, et al. (2004) Effects of treatment with anti-immunoglobulin E antibody omalizumab on airway inflammation in allergic asthma. Am J Respir Crit Care Med 170: 583–593. doi: 10.1164/rccm.200312-1651OC
|
| [32] |
Prussin C, Griffith DT, Boesel KM, et al. (2003) Omalizumab treatment downregulates dendritic cell FcepsilonRI expression. J Allergy Clin Immunol 112: 1147–1154. doi: 10.1016/j.jaci.2003.10.003
|
| [33] |
Huang YC, Leyko B, Frieri M (2005) Effects of omalizumab and budesonide on markers of inflammation in human bronchial epithelial cells. Ann Allergy Asthma Immunol 95: 443–451. doi: 10.1016/S1081-1206(10)61170-2
|
| [34] |
Gounni AS, Wellemans V, Yang J, et al. (2005) Human airway smooth muscle cells express the high affinity receptor for IgE (Fc epsilon RI): A critical role of Fc epsilon RI in human airway smooth muscle cell function. J Immunol 175: 2613–2621. doi: 10.4049/jimmunol.175.4.2613
|
| [35] |
Roth M, Tamm M (2010) The effects of omalizumab on IgE-induced cytokine synthesis by asthmatic airway smooth muscle cells. Ann Allergy Asthma Immunol 104: 152–160. doi: 10.1016/j.anai.2009.11.022
|
| [36] |
Hoshino M, Ohtawa J (2012) Effects of adding omalizumab, an anti-immunoglobulin E antibody, on airway wall thickening in asthma. Respiration 83: 520–528. doi: 10.1159/000334701
|
| [37] |
Tajiri T, Niimi A, Matsumoto H, et al. (2014) Comprehensive efficacy of omalizumab for severe refractory asthma: A time-series observational study. Ann Allergy Asthma Immunol 113: 470–475. doi: 10.1016/j.anai.2014.06.004
|
| [38] |
Saglani S, Payne DN, Zhu J, et al. (2007) Early detection of airway wall remodeling and eosinophilic inflammation in preschool wheezers. Am J Respir Crit Care Med 176: 858–864. doi: 10.1164/rccm.200702-212OC
|
| [39] |
Grainge CL, Lau LC, Ward JA, et al. (2011) Effect of bronchoconstriction on airway remodeling in asthma. N Engl J Med 364: 2006–2015. doi: 10.1056/NEJMoa1014350
|
| [40] |
Borkowski TA, Jouvin MH, Lin SY, et al. (2001) Minimal requirements for IgE-mediated regulation of surface Fc epsilon RI. J Immunol 167: 1290–1296. doi: 10.4049/jimmunol.167.3.1290
|
| [41] | Campbell AM, Vignola AM, Chanez P, et al. (1994) Low-affinity receptor for IgE on human bronchial epithelial cells in asthma. Immunology 82: 506–508. |
| [42] |
Palaniyandi S, Tomei E, Li Z, et al. (2011) CD23-dependent transcytosis of IgE and immune complex across the polarized human respiratory epithelial cells. J Immunol 186: 3484–3496. doi: 10.4049/jimmunol.1002146
|
| [43] |
Mauri P, Riccio AM, Rossi R, et al. (2014) Proteomics of bronchial biopsies: Galectin-3 as a predictive biomarker of airway remodelling modulation in omalizumab-treated severe asthma patients. Immunol Lett 162: 2–10. doi: 10.1016/j.imlet.2014.08.010
|
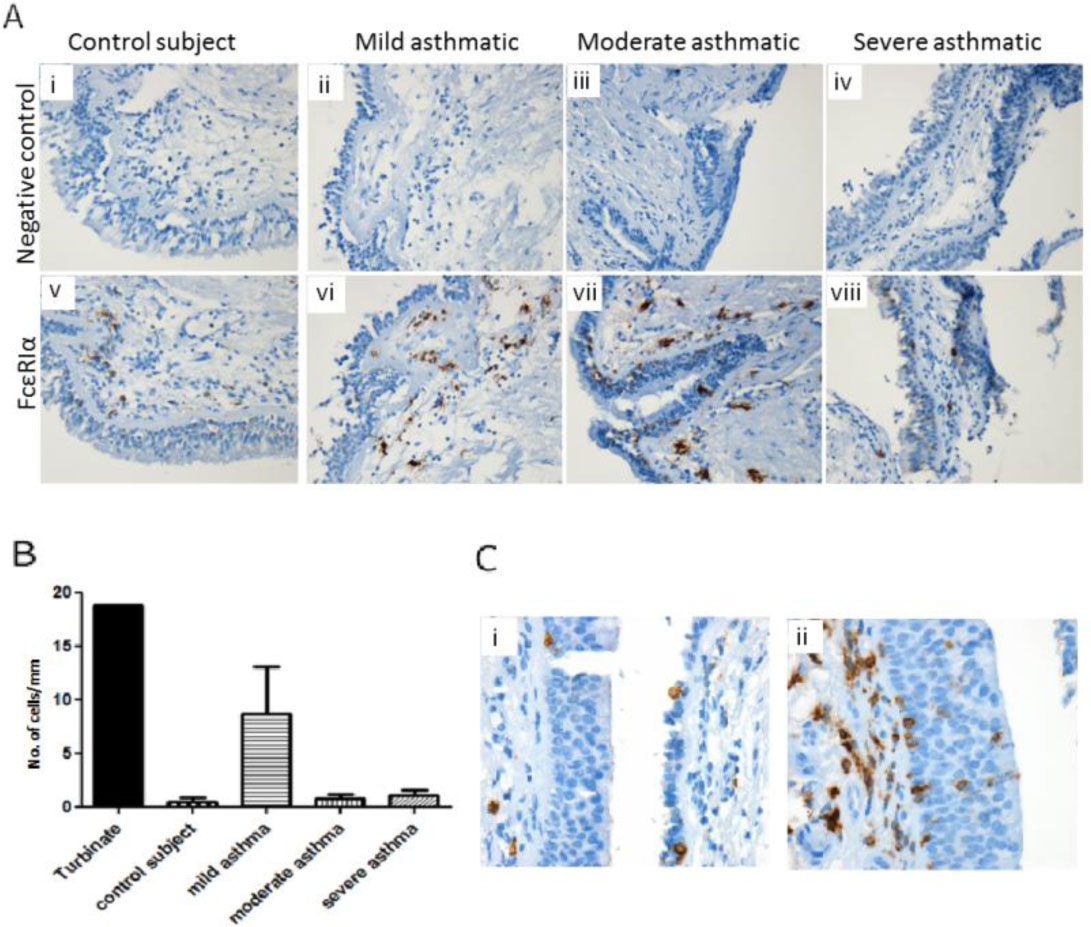
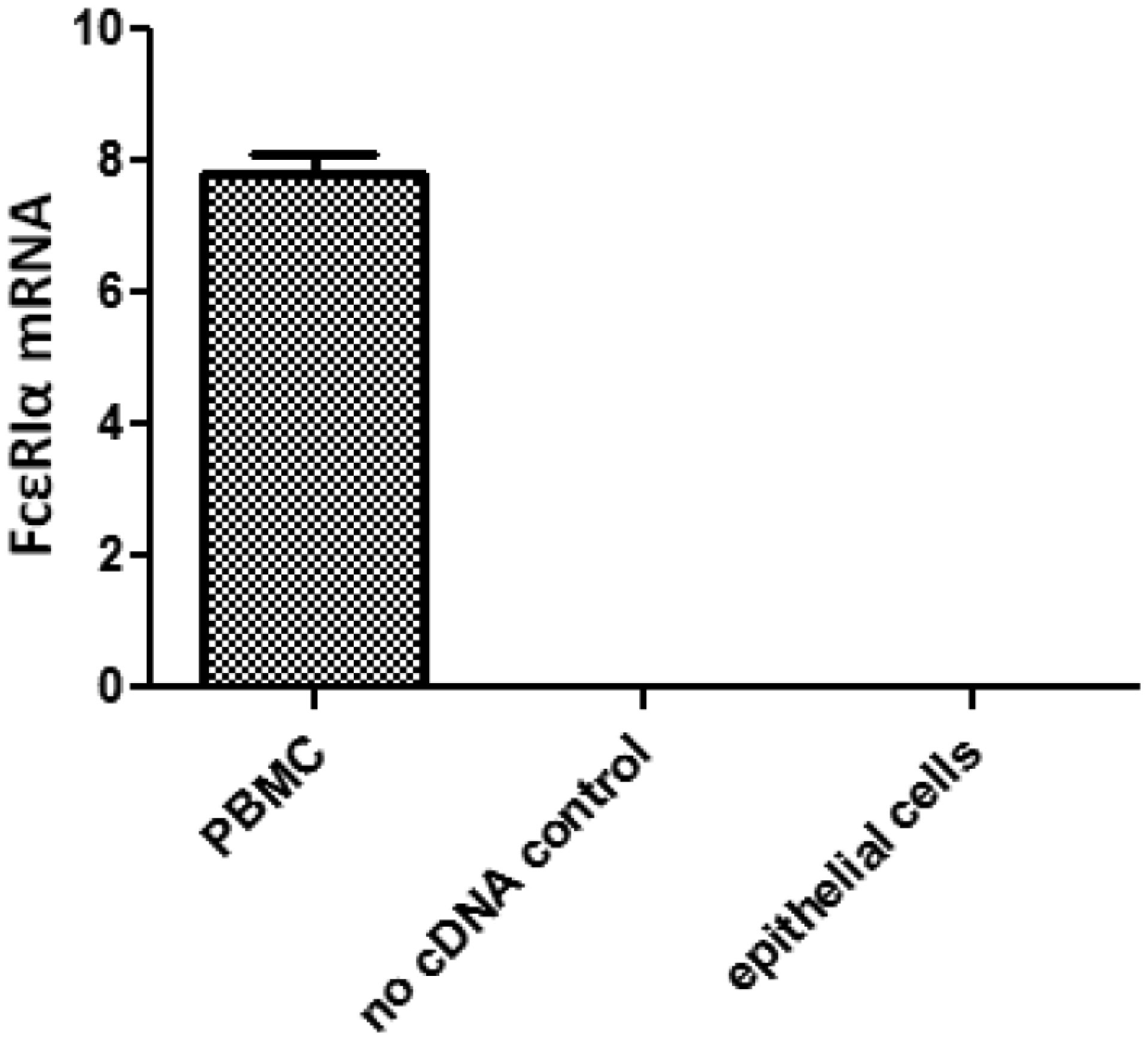
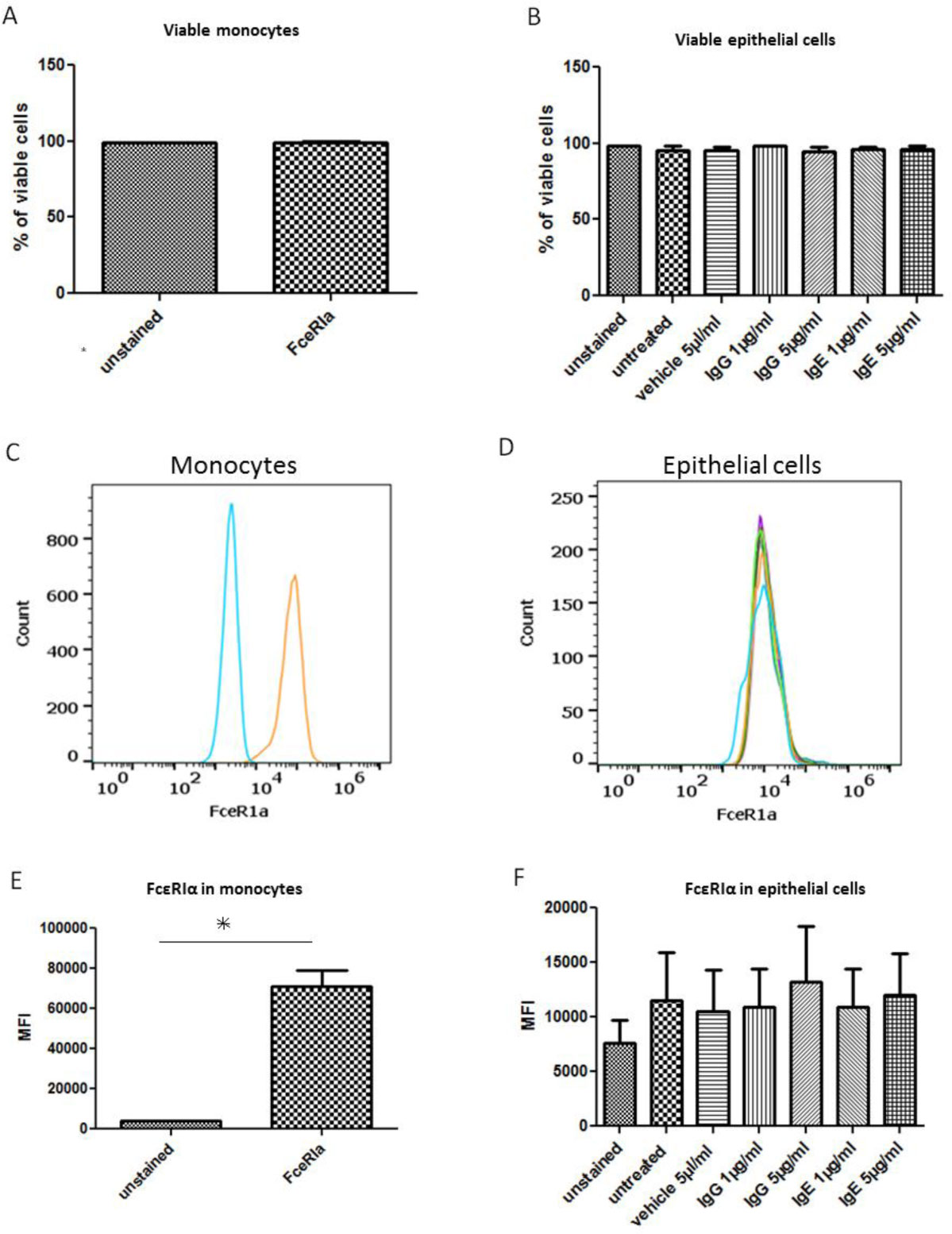
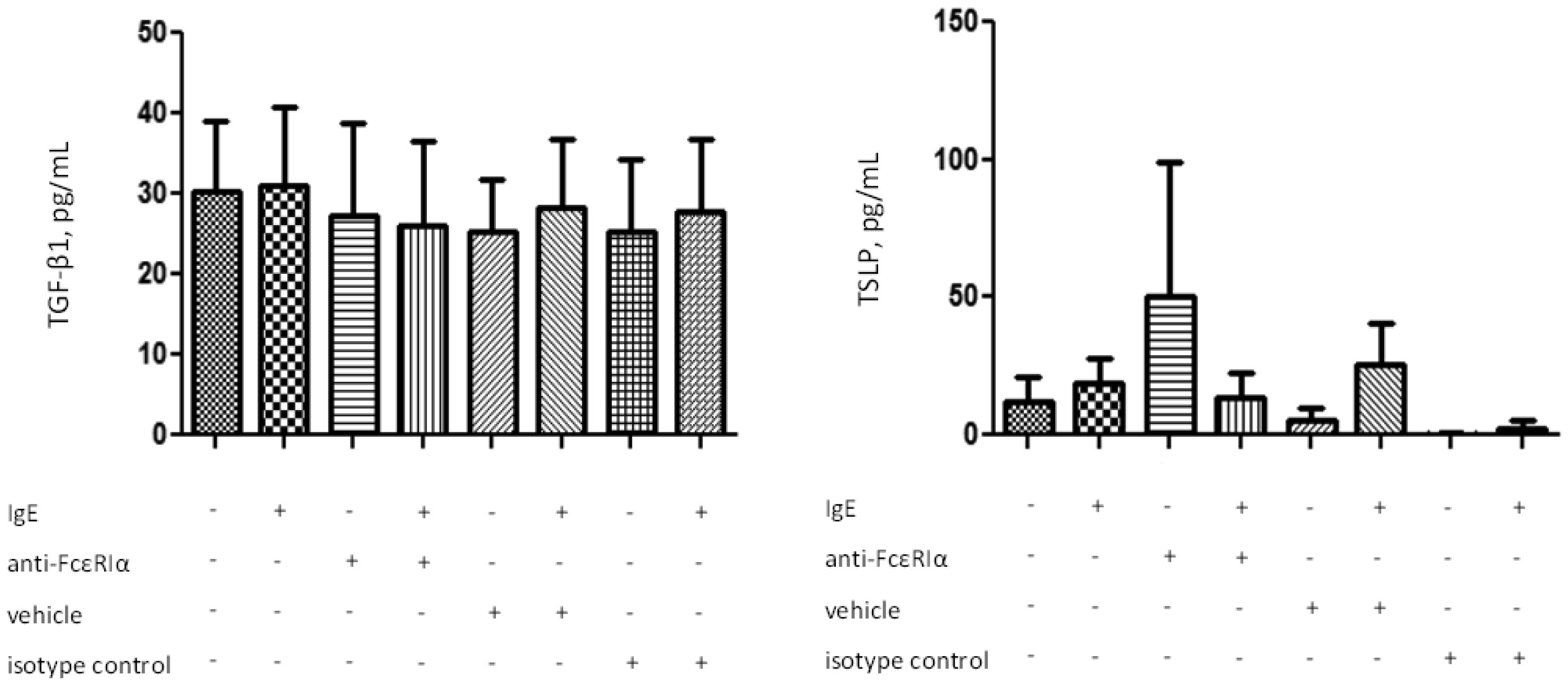
Figures(4) / Tables(2)

Omer Qibi, Severine Audusseau, Andrea Mogas, Zoulfia Allakhverdi, Abdelilah Soussi Gounni, Saba Al Heialy, Qutayba Hamid. No evidence for IgE receptor FcεRI expression on bronchial epithelial cells of asthmatic patients[J]. AIMS Allergy and Immunology, 2018, 2(4): 165-179. doi: 10.3934/Allergy.2018.4.165







 DownLoad:
DownLoad: