Citation: Chiara Schiraldi, Alberto D'Avino, Alessandro Ruggiero, Katia Della Corte, Mario De Rosa. Saccharomyces pastorianus as cell factory to improve production of fructose 1,6-diphosphate using novel fermentation strategies[J]. AIMS Bioengineering, 2015, 2(3): 206-221. doi: 10.3934/bioeng.2015.3.206

| [1] |
Markov AK, Warren ET, Cohly HHP, et al. (2007) Influence of Fructose-1,6-diphosphate on Endotoxin-Induced Lung Injuries in Sheep. J Surg Res 138: 45-50. doi: 10.1016/j.jss.2006.06.038

|
| [2] |
Sud V, Lindley SO, McDaniel O, et al. (2005) Evaluation of fructose 1,6-diphosphate for salvage of ischemic gracilis flaps in rats. J Reconstr Microsurg 21: 191-196. doi: 10.1055/s-2005-869826
|
| [3] | Zhou J, Wang F, Zhang J, et al. (2014) Repeated febrile convulsions impair hippocampal neurons and cause synaptic damage in immature rats: neuroprotective effect of fructose-1,6-diphosphate. NRR 9: 937-942. |
| [4] | Stringer JL, Xu K (2008) Possible mechanisms for the anticonvulsant activity of fructose-1,6-diphosphate. Epilepsia 49 Suppl 8:101-103. |
| [5] | Bisso GM, Melelli F (1986) Application of glutaraldehyde cross-linked yeast cells to continuous phosphorylation of glucose to fructose 1,6-diphosphate. Process Biochem 21:113-117. |
| [6] | Melelli F, Tommasi A, Bisso GM (1989) Time-course phosphorylation of glucose by Saccharomyces carlsbergensis following alterations in cell membrane permeability. Biotechnol Lett 2: 535-539. |
| [7] |
Compagno C, Tura A, Ranzi BM, et al. (1992) Production of fructose diphosphate by bioconversion of molasses with Saccharomyces cerevisiae cells. Biotechnol Lett 14: 495-498. doi: 10.1007/BF01023174
|
| [8] |
Compagno C, Tura A, Ranzi BM, et al. (1993) Bioconversion of lactose/whey to fructose diphosphate with recombinant Saccharomyces cerevisiae cells. Biotechnol Bioeng. 42: 398-400. doi: 10.1002/bit.260420319
|
| [9] | Widjaja A, Yasuda M, Ogino H, et al. (1999b) Enzymatic production of fructose 1,6-diphosphate with ATP regeneration in a batch reactor and a semi-batch reactor using crude cell extract of Bacillus stearothermophilus. J Biosci Bioeng 87: 693-696. |
| [10] | Shiroshima M, Widjaja A, Yasuda M, et al. (1999) Theoretical Investigation of Fructose 1,6-diphosphate Production and simultaneous ATP Regeneration by Conjugated Enzymes in an Ultrafiltration Hollow-Fiber Reactor. J Biosci Bioeng 88: 632-639 |
| [11] |
Schiraldi C, Adduci V, Valli V, et al. (2003). High cell density cultivation of probiotics and lactic acid production. Biotech Bioeng 82: 213-222. doi: 10.1002/bit.10557
|
| [12] |
Schiraldi C, Marulli F, Di Lernia I, et al. (1999) A microfiltration bioreactor to achieve high cell density in Sulfolobus solfataricus fermentation. Extremophiles 3: 199-204. doi: 10.1007/s007920050117
|
| [13] |
Alfano A, Donnarumma G, Cimini D, et al. (2015) Lactobacillus plantarum: Microfiltration Experiments for the Production of Probiotic Biomass to be Used in Food and Nutraceutical Preparations. Biotechnol Progr 31: 325-333. doi: 10.1002/btpr.2037
|
| [14] |
Schiraldi C, Acone M, Giuliano M, et al. (2001) Innovative fermentation strategies for the production of extremophilic enzymes. Extremophiles 5: 193-198. doi: 10.1007/s007920100194
|
| [15] | Schiraldi C, Martino A, Acone M, et al. (2000) Effective production of a thermostable alpha-glucosidase from Sulfolobus solfataricus in Escherichia coli exploiting a microfiltration bioreactor. Biotechnol Bioeng 70: 670-676. |
| [16] | Yu-Cai H, Feng L, Dan-Ping Z, et al. (2015) Biotransformation of 1,3-propanediol cyclic sulfate and its derivatives to diols by toluene-permeabilized cells of bacillus sp. CCZU11-1. Appl Biochem Biotechnol 175: 2647-2658. |
| [17] |
Lagunas R (1993) Sugar transport in Saccharomyces cerevisiae. FEMS Microbiol Rev 104: 229-242. doi: 10.1111/j.1574-6968.1993.tb05869.x
|
| [18] | Wills C (1996) Some puzzle about carbon catabolite repression in yeast. Res Microbiol 147:556-572. |
| [19] |
Kaeppeli O, Sonnleitner B (1986) Regulation of sugar metabolism in Saccharomyces-type yeast: experimental and conceptual considerations. Crit Rev Biotechnol 4: 299-325. doi: 10.3109/07388558609150798
|
| [20] |
Felix H (1982) Permeabilized cells. Anal Biochem 120: 211-234. doi: 10.1016/0003-2697(82)90340-2
|
| [21] |
Dipankar K, Allen JB (2010) Triton X-100 concentration effects on membrane permeability of a single HeLa cell by scanning electrochemical microscopy (SECM). Proc Natl Acad Sci U S A 107: 16783-16787. doi: 10.1073/pnas.1011614107
|
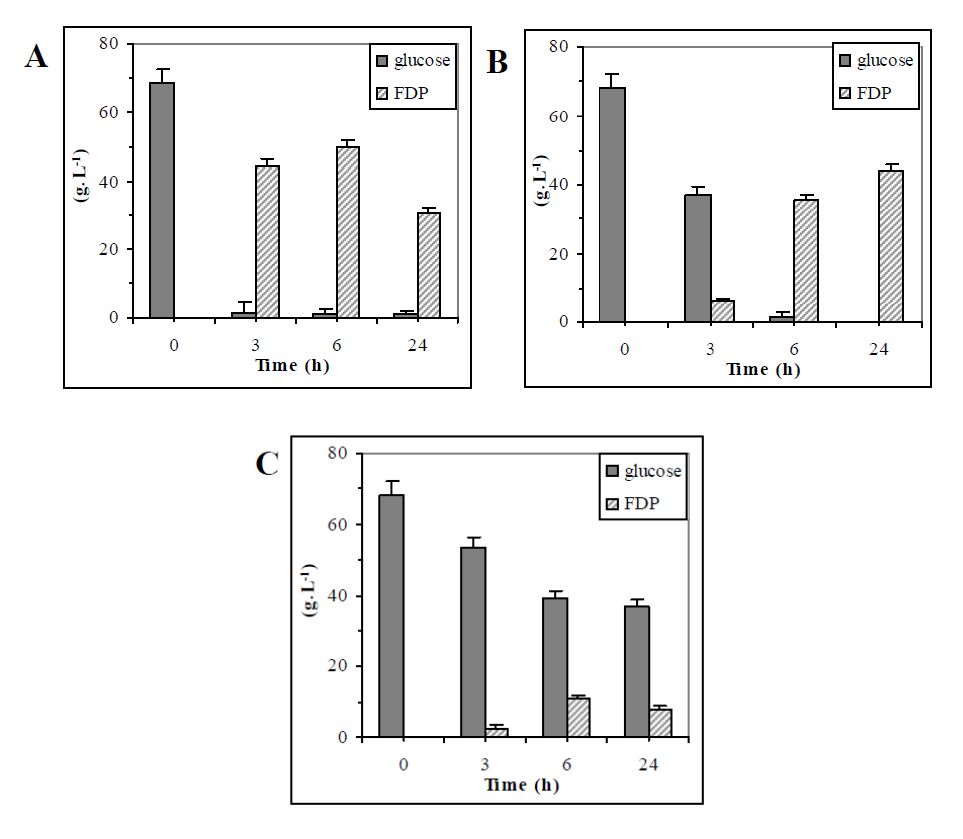
Figures(5) / Tables(2)

Chiara Schiraldi, Alberto D'Avino, Alessandro Ruggiero, Katia Della Corte, Mario De Rosa. Saccharomyces pastorianus as cell factory to improve production of fructose 1,6-diphosphate using novel fermentation strategies[J]. AIMS Bioengineering, 2015, 2(3): 206-221. doi: 10.3934/bioeng.2015.3.206







 DownLoad:
DownLoad: