Citation: Guang Long, Bo Lin, Lu Wang, Lingyan Wu, Tieying Yin, Donghong Yu, Guixue Wang. Sappan Lignum Extract Inhibits Restenosis in the Injured Artery through the Deactivation of Nuclear Factor-κB[J]. AIMS Bioengineering, 2014, 1(1): 25-39. doi: 10.3934/bioeng.2014.1.25

| [1] | Gaspardone A, Versaci F (2005) Coronary stenting and inflammation. Am J Cardiol 96:65-70. |
| [2] |
Monaco C, Paleolog E (2004) Nuclear factor kappaB: a potential therapeutic target in atherosclerosis and thrombosis. Cardiovasc Res 61: 671-682. doi: 10.1016/j.cardiores.2003.11.038

|
| [3] |
Yoshimura S, Morishita R, Hayashi K, et al. (2001) Inhibition of intimal hyperplasia after balloon injury in rat carotid artery model using cis-element 'decoy' of nuclear factor-kappaB binding site as a novel molecular strategy. Gene Ther 8: 1635-1642. doi: 10.1038/sj.gt.3301566
|
| [4] |
Kim EC, Hwang YS, Lee HJ, et al. (2005) Caesalpinia sappan induces cell death by increasing the expression of p53 and p21WAF1/CIP1 in head and neck cancer cells. Am J Chin Med 33: 405-414. doi: 10.1142/S0192415X05003016
|
| [5] |
Xu HX, Lee SF (2004) The antibacterial principle of Caesalpina sappan. Phytother Res 18:647-651. doi: 10.1002/ptr.1524
|
| [6] |
Sasaki Y, Hosokawa T, Nagai M, et al. (2007) In vitro study for inhibition of NO production about constituents of Sappan Lignum. Biol Pharm Bull 30: 193-196. doi: 10.1248/bpb.30.193
|
| [7] | Sasaki Y, Suzuki M, Matsumoto T, et al.(2010) Vasorelaxant activity of Sappan Lignum constituents and extracts on rat aorta and mesenteric artery. Biol Pharm Bull 33:1555-1560. |
| [8] |
Shen J, Zhang H, Lin H, et al. (2007) Brazilein protects the brain against focal cerebral ischemia reperfusion injury correlating to inflammatory response suppression. Eur J Pharmacol 558: 88-95. doi: 10.1016/j.ejphar.2006.11.059
|
| [9] |
Oh GT, Choi JH, Hong JJ, et al. (2001) Dietary hematein ameliorates fatty streak lesions in the rabbit by the possible mechanism of reducing VCAM-1 and MCP-1 expression. Atherosclerosis 159: 17-26. doi: 10.1016/S0021-9150(01)00464-6
|
| [10] |
Washiyama M, Sasaki Y, Hosokawa T, et al. (2009) Anti-inflammatory constituents of Sappan Lignum. Biol Pharm Bull 32: 941-944. doi: 10.1248/bpb.32.941
|
| [11] |
Wang X, Sun W, Sun H, et al. (2008) Analysis of the constituents in the rat plasma after oral administration of Yin Chen Hao Tang by UPLC/Q-TOF-MS/MS. J Pharm Biomed Anal 46: 477-490. doi: 10.1016/j.jpba.2007.11.014
|
| [12] |
Yang D, Cai S, Liu H, et al. (2006) On-line identification of the constituents of Buyang Huanwu decoction in pig serum using combined HPLC-DAD-MS techniques. J Chromatogr B Analyt Technol Biomed Life Sci 831: 288-302. doi: 10.1016/j.jchromb.2005.12.032
|
| [13] |
Matsuura K, Kawakita T, Nakai S, et al. (1993) Role of B-lymphocytes in the immunopharmacological effects of a traditional Chinese medicine, xiao-chai-hu-tang (shosaiko-to). Int J Immunopharmacol 15: 237-243. doi: 10.1016/0192-0561(93)90100-D
|
| [14] |
Umeda M, Amagaya S, Ogihara Y, et al. (1988) Effects of certain herbal medicines on the biotransformation of arachidonic acid: a new pharmacological testing method using serum. J Ethnopharmacol 23: 91-98. doi: 10.1016/0378-8741(88)90117-1
|
| [15] |
Ross R, Glomset J, Kariya B, et al. (1974) A platelet-dependent serum factor that stimulates the proliferation of arterial smooth muscle cells in vitro. Proc Natl Acad Sci 71:1207-1210. doi: 10.1073/pnas.71.4.1207
|
| [16] |
Lopez-Franco O, Suzuki Y, Sanjuan G, et al. (2002) Nuclear factor-kappa B inhibitors as potential novel anti-inflammatory agents for the treatment of immune glomerulonephritis. Am J Pathol 161: 1497-1505. doi: 10.1016/S0002-9440(10)64425-2
|
| [17] | Romano MF, Lamberti A, Tassone P, et al. (1998) Triggering of CD40 antigen inhibits fludarabine-induced apoptosis in B chronic lymphocytic leukemia cells. Blood 92:990-995. |
| [18] |
Massey V (1959) The microestimation of succinate and the extinction coefficient of cytochrome c. Biochim Biophys Acta 34: 255-256. doi: 10.1016/0006-3002(59)90259-8
|
| [19] | Yang TC, Zhang SW, Sun LN, et al. (2008) Ren AM. Magnolol attenuates sepsis-induced gastrointestinal dysmotility in rats by modulating inflammatory mediators. World J Gastroenterol 14: 7353-7360. |
| [20] |
Maier W, Altwegg LA, Corti R, et al. (2005) Inflammatory markers at the site of ruptured plaque in acute myocardial infarction: locally increased interleukin-6 and serum amyloid A but decreased C-reactive protein. Circulation 111: 1355-1361. doi: 10.1161/01.CIR.0000158479.58589.0A
|
| [21] |
Wung BS, Ni CW, Wang DL,et al. (2005) ICAM-1 induction by TNFalpha and IL-6 is mediated by distinct pathways via Rac in endothelial cells. J Biomed Sci 12: 91-101. doi: 10.1007/s11373-004-8170-z
|
| [22] |
Tanaka H, Sukhova G, Schwartz D, et al. (1996) Proliferating arterial smooth muscle cells after balloon injury express TNF-alpha but not interleukin-1 or basic fibroblast growth factor. Arterioscler Thromb Vasc Biol 16: 12-18. doi: 10.1161/01.ATV.16.1.12
|
| [23] |
Jovinge S, Hultgardh-Nilsson A, Regnstrom J, et al. (1997) Tumor necrosis factor-alpha activates smooth muscle cell migration in culture and is expressed in the balloon-injured rat aorta. Arterioscler Thromb Vasc Biol 17: 490-497. doi: 10.1161/01.ATV.17.3.490
|
| [24] |
Chen YH, Lin SJ, Chen JW, et al. (2002) Magnolol attenuates VCAM-1 expression in vitro in TNF-alpha-treated human aortic endothelial cells and in vivo in the aorta of cholesterol-fed rabbits. Br J Pharmacol 135: 37-47. doi: 10.1038/sj.bjp.0704458
|
| [25] |
Shrivastava A, Aggarwal BB (1999) Antioxidants differentially regulate activation of nuclear factor-kappa B, activator protein-1, c-jun amino-terminal kinases, and apoptosis induced by tumor necrosis factor: evidence that JNK and NF-kappa B activation are not linked to apoptosis. Antioxid Redox Signal 1: 181-191. doi: 10.1089/ars.1999.1.2-181
|
| [26] | Badami S, Moorkoth S, Rai SR, et al. (2003) Antioxidant activity of Caesalpinia sappan heartwood. Biol Pharm Bul l26: 1534-1537. |
| [27] | Choi BM, Lee JA, Gao SS, et al. (2007) Brazilin and the extract from Caesalpinia sappan L. protect oxidative injury through the expression of heme oxygenase-1. Biofactors 30:149-157 |
| [28] |
Lee JW, Lee BS, Lee JY, et al. (2011) The herbal extract HMC05 inhibits neointima formation in balloon-injured rat carotid arteries: possible therapeutic implications of HMC05. J Ethnopharmacol (Ireland) 133: 168-176. doi: 10.1016/j.jep.2010.09.015
|
| [29] |
Jeon J, Park KA, Lee H, et al. (2011) Water extract of Cynanchi atrati Radix regulates inflammation and apoptotic cell death through suppression of IKK-mediated NF-(ordM)B signalin. J Ethnopharmacol (Ireland) 137: 626-634. doi: 10.1016/j.jep.2011.06.022
|
| [30] | Matsui M, Adib-Conquy M, Coste A, et al. (2012) Aqueous extract of Vitex trifolia L. (Labiatae) inhibits LPS-dependent regulation of inflammatory mediators in RAW 264.7 macrophages through inhibition of Nuclear Factor kappa B translocation and expression. J Ethnopharmacol (Ireland) 143: 24-32. |
| [31] |
Shin W, Cuong TD, Lee JH, et al. (2011) Arginase Inhibition by Arginase Inhibition by Ethylacetate Extract of Caesalpinia sappan Lignum Contributes to Activation of Endothelial Nitric Oxide Synthase. Korean J Physiol Pharmacol 15: 123-128. doi: 10.4196/kjpp.2011.15.3.123
|
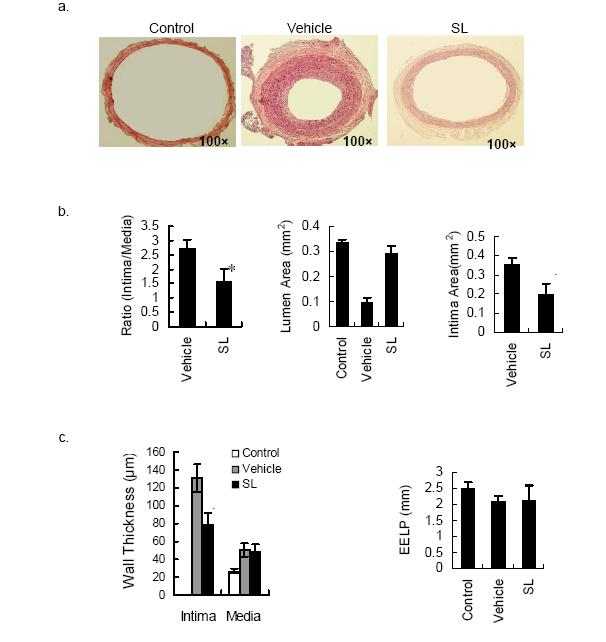
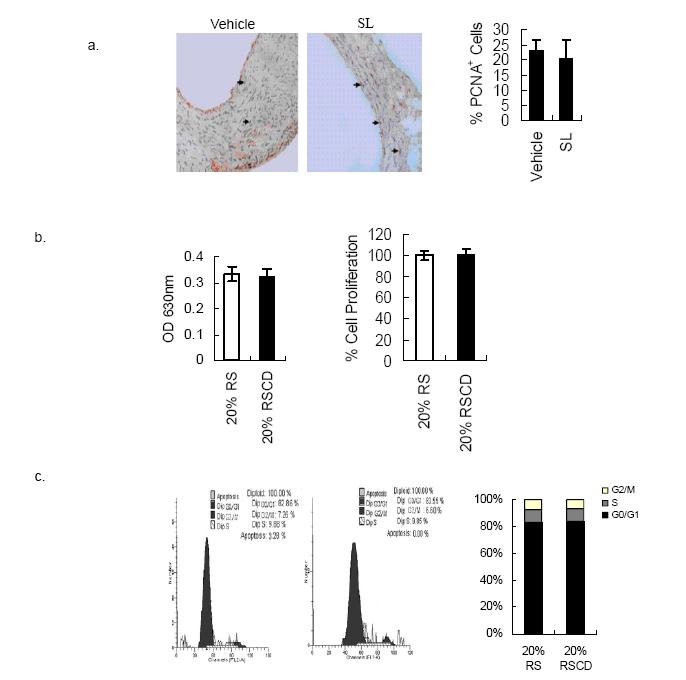
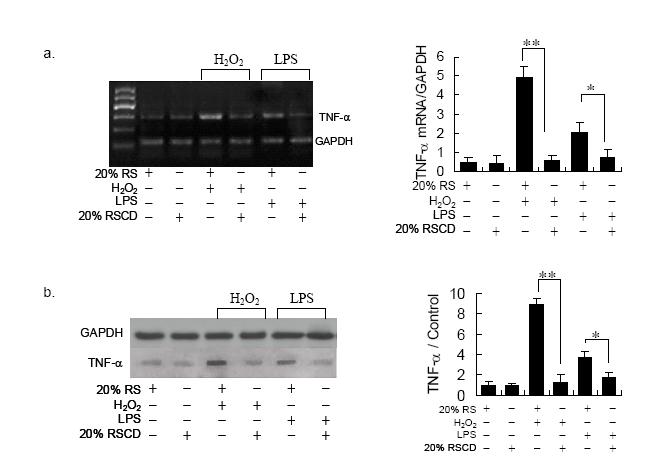
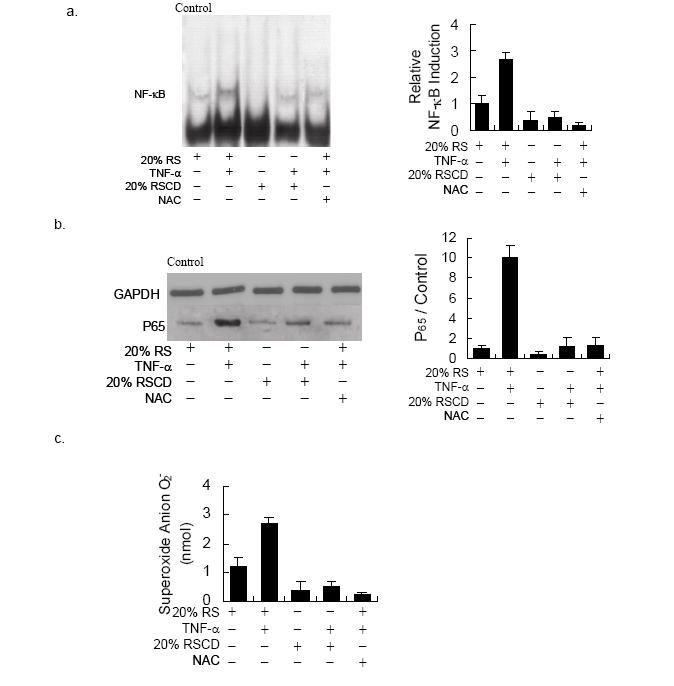
Figures(4)

Guang Long, Bo Lin, Lu Wang, Lingyan Wu, Tieying Yin, Donghong Yu, Guixue Wang. Sappan Lignum Extract Inhibits Restenosis in the Injured Artery through the Deactivation of Nuclear Factor-κB[J]. AIMS Bioengineering, 2014, 1(1): 25-39. doi: 10.3934/bioeng.2014.1.25







 DownLoad:
DownLoad: