Citation: Azhar Alhasawi, Vasu D. Appanna. Enhanced extracellular chitinase production in Pseudomonas fluorescens: biotechnological implications[J]. AIMS Bioengineering, 2017, 4(3): 366-375. doi: 10.3934/bioeng.2017.3.366

| [1] |
Khoushab F, Yamabhai M (2010) Chitin research revisited. Mar Drugs 8: 1988–2012. doi: 10.3390/md8071988

|
| [2] | Cheba BA (2011) Chitin and chitosan: marine biopolymers with unique properties and versatile applications. Global J Biotech Biochem 6: 149–153. |
| [3] | Thiagarajan V, Revathi R, Aparanjini K, et al. (2011) Extra cellular chitinase production by Streptomyces sp.PTK19 in submerged fermentation and its lytic activity on Fusarium oxysporum PTK2 cell wall. Int J Curr Sci 1: 30–44. |
| [4] |
Dahiya N, Tewari R, Hoondal GS (2006) Biotechnological aspects of chitinolytic enzymes: a review. Appl Microbiol Biot 71: 773–782. doi: 10.1007/s00253-005-0183-7
|
| [5] |
Haggag WM, Abdallh E (2012) Purification and characterization of chitinase produced by Endophytic ptomyceshygroscopicus against some phytopathogens. J Microbiol Res 2: 145–151. doi: 10.5923/j.microbiology.20120205.06
|
| [6] | Reissig JL, Strominger JL, Leloir LF (1955) A modified colorimetric method for the estimation of N-acetylamino sugars. J Biol Chem 21: 959–966. |
| [7] | Kamil Z, Saleh M, Moustafa S (2007) Isolation and identification of rhizosphere soil chitinolytic bacteria and their potential in antifungal biocontrol 1. Global J Mol Sci 2: 57–66. |
| [8] |
Bhattacharya D, Nagpure A, Gupta RK (2007) Bacterial chitinases: properties and potential. Crit Rev Biotechnol 27: 21–28. doi: 10.1080/07388550601168223
|
| [9] | Hamid R, Khan MA, Ahmad M, et al. (2013) Chitinases: an update. J Pharm Bioallied Sci 5: 21–29. |
| [10] | Narayanan K, Chopade N, Raj PV, et al. (2013) Fungal chitinase production and its application in biowaste management. J Sci Ind Res 72: 393–399. |
| [11] | Appanna VD, Pierre MS (1996) Aluminum elicits exocellular phosphatidylethanolamine production in Pseudomonas fluorescens. Appl Environ Microb 62: 2778–2782. |
| [12] |
Auger C, Han S, Appanna VP, et al. (2013) Metabolic reengineering invoked by microbial systems to decontaminate aluminum: implications for bioremediation technologies. Biotechnol Adv 31: 266–273. doi: 10.1016/j.biotechadv.2012.11.008
|
| [13] |
Appanna VD, Gazsó LG, Pierre MS (1996) Multiple-metal tolerance in Pseudomonas fluorescens and its biotechnological significance. J Biotechnol 52: 75–80. doi: 10.1016/S0168-1656(96)01623-9
|
| [14] |
Bignucolo A, Appanna VP, Thomas SC, et al. (2013) Hydrogen peroxide stress provokes a metabolic reprogramming in Pseudomonas fluorescens: enhanced production of pyruvate. J Biotechnol 167: 309–315. doi: 10.1016/j.jbiotec.2013.07.002
|
| [15] |
Middaugh J, Hamel R, Jean-Baptiste G, et al. (2005) Aluminum triggers decreased aconitase activity via Fe-S cluster disruption and the overexpression of isocitrate dehydrogenase and isocitrate lyase: a metabolic network mediating cellular survival. J Biol Chem 280: 3159–3165. doi: 10.1074/jbc.M411979200
|
| [16] | Yanes ML, Bajsa N (2016) Fluorescent Pseudomonas: a natural resource from soil to enhance crop growth and health, In: Microbial models: from environmental to industrial sustainability Springer Singapore, 323–349. |
| [17] |
Chenier D, Beriault R, Mailloux R, et al. (2008) Involvement of fumarase C and NADH oxidase in metabolic adaptation of Pseudomonas fluorescens cells evoked by aluminum and gallium toxicity. Appl Environ Microb 74: 3977–3984. doi: 10.1128/AEM.02702-07
|
| [18] |
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248–254. doi: 10.1016/0003-2697(76)90527-3
|
| [19] |
Mailloux R, Lemire J, Kalyuzhnyi S, et al. (2008) A novel metabolic network leads to enhanced citrate biogenesis in Pseudomonas fluorescens exposed to aluminum toxicity. Extremophiles 12: 451–459. doi: 10.1007/s00792-008-0150-1
|
| [20] |
Alhasawi A, Auger C, Appanna VP, et al. (2014) Zinc toxicity and ATP production in Pseudomonas fluorescens. J Appl Microbial 117: 65–73. doi: 10.1111/jam.12497
|
| [21] |
Sharp RG (2013) A review of the applications of chitin and its derivatives in agriculture to modify plant-microbial interactions and improve crop yields. Agronomy 3: 757–793. doi: 10.3390/agronomy3040757
|
| [22] |
Neiendam NM, Sørensen J (1999) Chitinolytic activity of Pseudomonas fluorescens isolates from barley and sugar beet rhizosphere. FEMS Microbiol Ecol 30: 217–227. doi: 10.1111/j.1574-6941.1999.tb00650.x
|
| [23] |
Negrulescu A, Patrulea V, Mincea MM, et al. (2012) Adapting the reducing sugars method with dinitrosalicylic acid to microtiter plates and microwave heating. J Braz Chem Soc 23: 2176–2182. doi: 10.1590/S0103-50532013005000003
|
| [24] |
Auger C, Lemire J, Cecchini D, et al. (2011) The metabolic reprogramming evoked by nitrosative stress triggers the anaerobic utilization of citrate in Pseudomonas fluorescens. PloS One 6: e28469. doi: 10.1371/journal.pone.0028469
|
| [25] | Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 277: 680–685. |
| [26] |
Liu CL, Shen CR, Hsu FF, et al. (2009) Isolation and identification of two novel SDS‐resistant secreted chitinases from Aeromonas schubertii. Biotechnol Prog 25: 124–131. doi: 10.1002/btpr.100
|
| [27] |
Loni PP, Patil JU, Phugare SS, et al. (2014) Purification and characterization of alkaline chitinase from Paenibacillus pasadenensis NCIM 5434. J Basic Microb 54: 1080–1089. doi: 10.1002/jobm.201300533
|
| [28] | Velusamy P, Ko HS, Kim KY (2011) Determination of antifungal activity of Pseudomonas sp. A3 against Fusarium oxysporum by high performance liquid chromatography (HPLC). Agric Food Annal Bacteriol 1: 15–23. |
| [29] |
Zou X, Nonogaki H, Welbaum GE (2002) A gel diffusion assay for visualization and quantification of chitinase activity. Mol Biotechnol 22: 19–23. doi: 10.1385/MB:22:1:019
|
| [30] | Karunya SK (2011) Optimization and purification of chitinase produced by Bacillus subtilis and its antifungal activity against plant pathogens. Intl J Pharma Biol Arch 2: 1680–1685. |
| [31] | Viarsagh MS, Janmaleki M, Falahatpisheh HR, et al. (2010) Chitosan preparation from Persian Gulf shrimp shells and investigating the effect of time on the degree of deacetylation. J Paramed Sci 1: 2–6. |
| [32] | Kuzu SB, Güvenmez HK, Denizci AA (2012) Production of a thermostable and alkaline chitinase by Bacillus thuringiensis subsp. kurstaki strain HBK-51. Biotechnol Res Int 2012: 1–6. |
| [33] | Park SH, Lee JH, Lee HK (2000) Purification and characterization of chitinase from a marine bacterium, Vibrio sp. 98CJ11027. J Microbiol 38: 224–229. |
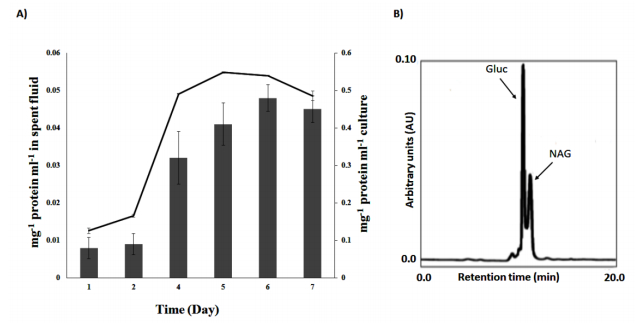
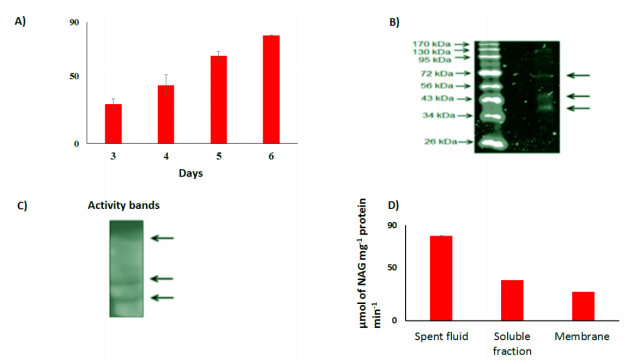
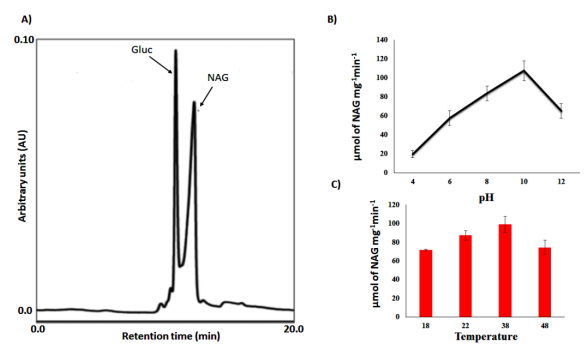
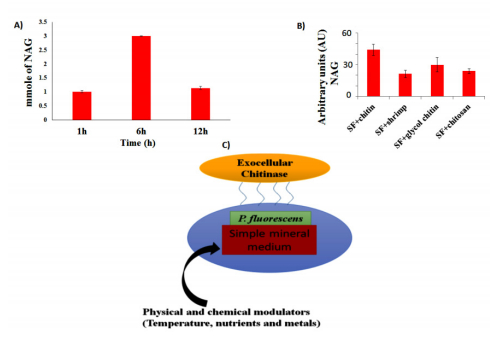
Figures(4) / Tables(1)

Azhar Alhasawi, Vasu D. Appanna. Enhanced extracellular chitinase production in Pseudomonas fluorescens: biotechnological implications[J]. AIMS Bioengineering, 2017, 4(3): 366-375. doi: 10.3934/bioeng.2017.3.366







 DownLoad:
DownLoad: