Citation: Renata Tisi, Marco Rigamonti, Silvia Groppi, Fiorella Belotti. Calcium homeostasis and signaling in fungi and their relevance for pathogenicity of yeasts and filamentous fungi[J]. AIMS Molecular Science, 2016, 3(4): 505-549. doi: 10.3934/molsci.2016.4.505

| [1] |
Wolfe D, Pearce D (2006) Channeling studies in yeast: yeast as a model for channelopathies? Neuromolecular Med 8: 279-306. doi: 10.1385/NMM:8:3:279

|
| [2] |
Cui J, Kaandorp J, Ositelu O, et al. (2009) Simulating calcium influx and free calcium concentrations in yeast. Cell Calcium 45: 123-132. doi: 10.1016/j.ceca.2008.07.005
|
| [3] |
Ton V, Rao R (2004) Functional expression of heterologous proteins in yeast: insights into Ca2+ signaling and Ca2+-transporting ATPases. Am J Physiol Cell Physiol 287: C580-589. doi: 10.1152/ajpcell.00135.2004
|
| [4] | Iida H, Yagawa Y, Anraku Y (1990) Essential role for induced Ca2+ influx followed by [Ca2+]i rise in maintaining viability of yeast cells late in the mating pheromone response pathway. A study of [Ca2+]i in single Saccharomyces cerevisiae cells with imaging of fura-2. J Biol Chem 265: 13391-13399. |
| [5] | Halachmi D, Eilam Y (1993) Calcium homeostasis in yeast cells exposed to high concentrations of calcium. Roles of vacuolar H+-ATPase and cellular ATP. FEBS Lett 316: 73-78. |
| [6] | Berridge MJ, Lipp P, Bootman MD (2000) The versatility and universality of calcium signalling. Nat Rev Mol Cell Biol 1, 11-21. |
| [7] | Dunn T, Gable K, Beeler T (1994) Regulation of cellular Ca2+ by yeast vacuoles. J Biol Chem 269: 7273-7278. |
| [8] |
Zelter A, Bencina M, Bowman B, et al. (2004) A comparative genomic analysis of the calcium signaling machinery in Neurospora crassa, Magnaporthe grisea, and Saccharomyces cerevisiae. Fungal Genet Biol 41: 827-841. doi: 10.1016/j.fgb.2004.05.001
|
| [9] |
Bencina M, Bagar T, Lah L, et al. (2009) A comparative genomic analysis of calcium and proton signaling/homeostasis in Aspergillus species. Fungal Genet Biol 46: S93-S104. doi: 10.1016/j.fgb.2008.07.019
|
| [10] |
Pittman JK (2011) Vacuolar Ca2+ uptake. Cell Calcium 50: 139-146. doi: 10.1016/j.ceca.2011.01.004
|
| [11] | Brini M, Cali T, Ottolini D, et al. (2012) Calcium pumps: why so many? Compr Physiol 2: 1045-1060. |
| [12] |
Palmgren MG, Axelsen KB (1998) Evolution of P-type ATPases. Biochim Biophys Acta 1365: 37-45. doi: 10.1016/S0005-2728(98)00041-3
|
| [13] | Vandecaetsbeek I, Vangheluwe P, Raeymaekers L, et al. (2011) The Ca2+ pumps of the endoplasmic reticulum and Golgi apparatus. Cold Spring Harb Perspect Biol 3: 725-738. |
| [14] |
He W, Hu Z (2012) The role of the Golgi-resident SPCA Ca(2)+/Mn(2)+ pump in ionic homeostasis and neural function. Neurochem Res 37: 455-468. doi: 10.1007/s11064-011-0644-6
|
| [15] |
Brini M, Cali T, Ottolini D, et al. (2013) The plasma membrane calcium pump in health and disease. FEBS J 280: 5385-5397. doi: 10.1111/febs.12193
|
| [16] |
Brini M, Carafoli E (2009) Calcium pumps in health and disease. Physiol Rev 89: 1341-1378. doi: 10.1152/physrev.00032.2008
|
| [17] | Cunningham K, Fink G (1994) Ca2+ transport in Saccharomyces cerevisiae. J Exp Biol 196: 157-166. |
| [18] |
Bowman BJ, Draskovic M, Freitag M, et al. (2009) Structure and distribution of organelles and cellular location of calcium transporters in Neurospora crassa. Eukaryot Cell 8: 1845-1855. doi: 10.1128/EC.00174-09
|
| [19] |
Bowman BJ, Abreu S, Margolles-Clark E, et al. (2011) Role of four calcium transport proteins, encoded by nca-1, nca-2, nca-3, and cax, in maintaining intracellular calcium levels in Neurospora crassa. Eukaryot Cell 10: 654-661. doi: 10.1128/EC.00239-10
|
| [20] |
Stathopoulos A, Cyert M (1997) Calcineurin acts through the CRZ1/TCN1-encoded transcription factor to regulate gene expression in yeast. Genes Dev 11: 3432-3444. doi: 10.1101/gad.11.24.3432
|
| [21] |
Matheos D, Kingsbury T, Ahsan U, et al. (1997) Tcn1p/Crz1p, a calcineurin-dependent transcription factor that differentially regulates gene expression in Saccharomyces cerevisiae. Genes Dev 11: 3445-3458. doi: 10.1101/gad.11.24.3445
|
| [22] |
Cunningham K, Fink G (1994) Calcineurin-dependent growth control in Saccharomyces cerevisiae mutants lacking PMC1, a homolog of plasma membrane Ca2+ ATPases. J Cell Biol 124: 351-363. doi: 10.1083/jcb.124.3.351
|
| [23] |
Findon H, Calcagno-Pizarelli AM, Martinez JL, et al. (2010) Analysis of a novel calcium auxotrophy in Aspergillus nidulans. Fungal Genet Biol 47: 647-655. doi: 10.1016/j.fgb.2010.04.002
|
| [24] |
Dinamarco TM, Freitas FZ, Almeida RS, et al. (2012) Functional characterization of an Aspergillus fumigatus calcium transporter (PmcA) that is essential for fungal infection. PLoS One 7: e37591. doi: 10.1371/journal.pone.0037591
|
| [25] |
Jiang H, Liu F, Zhang S, et al. (2014) Putative PmrA and PmcA are important for normal growth, morphogenesis and cell wall integrity, but not for viability in Aspergillus nidulans. Microbiology 160: 2387-2395. doi: 10.1099/mic.0.080119-0
|
| [26] |
Fan W, Idnurm A, Breger J, et al. (2007) Eca1, a sarcoplasmic/endoplasmic reticulum Ca2+-ATPase, is involved in stress tolerance and virulence in Cryptococcus neoformans. Infect Immun 75: 3394-3405. doi: 10.1128/IAI.01977-06
|
| [27] |
Adamikova L, Straube A, Schulz I, et al. (2004) Calcium signaling is involved in dynein-dependent microtubule organization. Mol Biol Cell 15: 1969-1980. doi: 10.1091/mbc.E03-09-0675
|
| [28] |
Rudolph H, Antebi A, Fink G, et al. (1989) The yeast secretory pathway is perturbed by mutations in PMR1, a member of a Ca2+ ATPase family. Cell 58: 133-145. doi: 10.1016/0092-8674(89)90410-8
|
| [29] |
Antebi A, Fink G (1992) The yeast Ca2+-ATPase homologue, PMR1, is required for normal Golgi function and localizes in a novel Golgi-like distribution. Mol Biol Cell 3: 633-654. doi: 10.1091/mbc.3.6.633
|
| [30] |
Halachmi D, Eilam Y (1996) Elevated cytosolic free Ca2+ concentrations and massive Ca2+ accumulation within vacuoles, in yeast mutant lacking PMR1, a homolog of Ca2+ -ATPase. FEBS Lett 392: 194-200. doi: 10.1016/0014-5793(96)00799-5
|
| [31] |
Strayle, J, Pozzan T, Rudolph H (1999) Steady-state free Ca2+ in the yeast endoplasmic reticulum reaches only 10 microM and is mainly controlled by the secretory pathway pump pmr1. EMBO J 18: 4733-4743. doi: 10.1093/emboj/18.17.4733
|
| [32] |
D��rr G, Strayle J, Plemper R, et al. (1998) The medial-Golgi ion pump Pmr1 supplies the yeast secretory pathway with Ca2+ and Mn2+ required for glycosylation, sorting, and endoplasmic reticulum-associated protein degradation. Mol Biol Cell 9: 1149-1162. doi: 10.1091/mbc.9.5.1149
|
| [33] |
Hu Z, Bonifas JM, Beech J, et al. (2000) Mutations in ATP2C1, encoding a calcium pump, cause Hailey-Hailey disease. Nat Genet 24: 61-65. doi: 10.1038/71701
|
| [34] |
Behne MJ, Tu CL, Aronchik I, et al. (2003) Human keratinocyte ATP2C1 localizes to the Golgi and controls Golgi Ca2+ stores. J Invest Dermatol 121: 688-694. doi: 10.1046/j.1523-1747.2003.12528.x
|
| [35] |
Cronin S, Rao R, Hampton R (2002) Cod1p/Spf1p is a P-type ATPase involved in ER function and Ca2+ homeostasis. J Cell Biol 157: 1017-1028. doi: 10.1083/jcb.200203052
|
| [36] |
Cronin S, Khoury A, Ferry D, et al. (2000) Regulation of HMG-CoA reductase degradation requires the P-type ATPase Cod1p/Spf1p. J Cell Biol 148: 915-924. doi: 10.1083/jcb.148.5.915
|
| [37] |
Vashist S, Frank CG, Jakob CA, et al. (2002) Two distinctly localized p-type ATPases collaborate to maintain organelle homeostasis required for glycoprotein processing and quality control. Mol Biol Cell 13: 3955-3966. doi: 10.1091/mbc.02-06-0090
|
| [38] |
Yu Q, Wang H, Xu N, et al. (2012) Spf1 strongly influences calcium homeostasis, hyphal development, biofilm formation and virulence in Candida albicans. Microbiology 158: 2272-2282. doi: 10.1099/mic.0.057232-0
|
| [39] |
Yu Q, Ding X, Zhang B, et al. (2013) The P-type ATPase Spf1 is required for endoplasmic reticulum functions and cell wall integrity in Candida albicans. Int J Med Microbiol 303: 257-266. doi: 10.1016/j.ijmm.2013.05.003
|
| [40] |
Krumpe K, Frumkin I, Herzig Y, et al. (2012) Ergosterol content specifies targeting of tail-anchored proteins to mitochondrial outer membranes. Mol Biol Cell 23: 3927-3935. doi: 10.1091/mbc.E11-12-0994
|
| [41] |
Cohen Y, Megyeri M, Chen OC, et al. (2013) The yeast p5 type ATPase, spf1, regulates manganese transport into the endoplasmic reticulum. PLoS One 8: e85519. doi: 10.1371/journal.pone.0085519
|
| [42] |
Miseta A, Kellermayer R, Aiello D, et al. (1999) The vacuolar Ca2+/H+ exchanger Vcx1p/Hum1p tightly controls cytosolic Ca2+ levels in S. cerevisiae. FEBS Lett 451: 132-136. doi: 10.1016/S0014-5793(99)00519-0
|
| [43] |
Cai X, Lytton J (2004) The cation/Ca2+ exchanger superfamily: phylogenetic analysis and structural implications. Mol Biol Evol 21: 1692-1703. doi: 10.1093/molbev/msh177
|
| [44] |
Shigaki T, Rees I, Nakhleh L, et al. (2006) Identification of three distinct phylogenetic groups of CAX cation/proton antiporters. J Mol Evol 63: 815-825. doi: 10.1007/s00239-006-0048-4
|
| [45] |
Pozos T, Sekler I, Cyert M (1996) The product of HUM1, a novel yeast gene, is required for vacuolar Ca2+/H+ exchange and is related to mammalian Na+/Ca2+ exchangers. Mol Cell Biol 16: 3730-3741. doi: 10.1128/MCB.16.7.3730
|
| [46] | Forster C, Kane P (2000) Cytosolic Ca2+ homeostasis is a constitutive function of the V-ATPase in Saccharomyces cerevisiae. J Biol Chem 275: 38245-38253. |
| [47] |
Waight AB, Pedersen BP, Schlessinger A, et al. (2013) Structural basis for alternating access of a eukaryotic calcium/proton exchanger. Nature 499: 107-110. doi: 10.1038/nature12233
|
| [48] | Kingsbury TJ, Cunningham KW (2000) A conserved family of calcineurin regulators. Genes Dev 14: 1595-1604. |
| [49] |
Spielvogel A, Findon H, Arst HN, et al. (2008) Two zinc finger transcription factors, CrzA and SltA, are involved in cation homoeostasis and detoxification in Aspergillus nidulans. Biochem J 414: 419-429. doi: 10.1042/BJ20080344
|
| [50] |
Kmetzsch L, Staats CC, Simon E, et al. (2010) The vacuolar Ca2+ exchanger Vcx1 is involved in calcineurin-dependent Ca2+ tolerance and virulence in Cryptococcus neoformans. Eukaryot Cell 9: 1798-1805. doi: 10.1128/EC.00114-10
|
| [51] |
Cagnac O, Aranda-Sicilia MN, Leterrier M, et al. (2010) Vacuolar cation/H+ antiporters of Saccharomyces cerevisiae. J Biol Chem 285: 33914-33922. doi: 10.1074/jbc.M110.116590
|
| [52] |
Demaegd D, Foulquier F, Colinet AS, et al. (2013) Newly characterized Golgi-localized family of proteins is involved in calcium and pH homeostasis in yeast and human cells. Proc Natl Acad Sci U S A 110: 6859-6864. doi: 10.1073/pnas.1219871110
|
| [53] |
Foulquier F, Amyere M, Jaeken J, et al. (2012) TMEM165 deficiency causes a congenital disorder of glycosylation. Am J Hum Genet 91: 15-26. doi: 10.1016/j.ajhg.2012.05.002
|
| [54] |
Cagnac O, Leterrier M, Yeager M, et al. (2007) Identification and characterization of Vnx1p, a novel type of vacuolar monovalent cation/H+ antiporter of Saccharomyces cerevisiae. J Biol Chem 282: 24284-24293. doi: 10.1074/jbc.M703116200
|
| [55] |
Manohar M, Mei H, Franklin AJ, et al. (2010) Zebrafish (Danio rerio) endomembrane antiporter similar to a yeast cation/H+ transporter is required for neural crest development. Biochemistry 49: 6557-6566. doi: 10.1021/bi100362k
|
| [56] |
Hong S, Cong X, Jing H, et al. (2013) Characterization of Ca2+/H+ exchange in the plasma membrane of Saccharomyces cerevisiae. Arch Biochem Biophys 537: 125-132. doi: 10.1016/j.abb.2013.07.005
|
| [57] |
Fischer M, Schnell N, Chattaway J, et al. (1997) The Saccharomyces cerevisiae CCH1 gene is involved in calcium influx and mating. FEBS Lett 419: 259-262. doi: 10.1016/S0014-5793(97)01466-X
|
| [58] |
Iida H, Nakamura H, Ono T, et al. (1994) MID1, a novel Saccharomyces cerevisiae gene encoding a plasma membrane protein, is required for Ca2+ influx and mating. Mol Cell Biol 14: 8259-8271. doi: 10.1128/MCB.14.12.8259
|
| [59] |
Martin DC, Kim H, Mackin NA, et al. (2011) New regulators of a high affinity Ca2+ influx system revealed through a genome-wide screen in yeast. J Biol Chem 286: 10744-10754. doi: 10.1074/jbc.M110.177451
|
| [60] |
Ding X, Yu Q, Xu N, et al. (2013) Ecm7, a regulator of HACS, functions in calcium homeostasis maintenance, oxidative stress response and hyphal development in Candida albicans. Fungal Genet Biol 57: 23-32. doi: 10.1016/j.fgb.2013.05.010
|
| [61] |
Tasaka Y, Nakagawa Y, Sato C, et al. (2000) yam8+, a Schizosaccharomyces pombe gene, is a potential homologue of the Saccharomyces cerevisiae MID1 gene encoding a stretch-activated Ca2+-permeable channel. Biochem Biophys Res Commun 269: 265-269. doi: 10.1006/bbrc.2000.2278
|
| [62] |
Lew RR, Abbas Z, Anderca MI, et al. (2008) Phenotype of a mechanosensitive channel mutant, mid-1, in a filamentous fungus, Neurospora crassa. Eukaryot Cell 7: 647-655. doi: 10.1128/EC.00411-07
|
| [63] |
Wang S, Cao J, Liu X, et al. (2012) Putative calcium channels CchA and MidA play the important roles in conidiation, hyphal polarity and cell wall components in Aspergillus nidulans. PLoS One 7: e46564. doi: 10.1371/journal.pone.0046564
|
| [64] |
Brand A, Lee K, Veses V, et al. (2009) Calcium homeostasis is required for contact-dependent helical and sinusoidal tip growth in Candida albicans hyphae. Mol Microbiol 71: 1155-1164. doi: 10.1111/j.1365-2958.2008.06592.x
|
| [65] |
Liu M, Du P, Heinrich G, et al. (2006) Cch1 mediates calcium entry in Cryptococcus neoformans and is essential in low-calcium environments. Eukaryot Cell 5: 1788-1796. doi: 10.1128/EC.00158-06
|
| [66] |
Cavinder B, Hamam A, Lew RR, et al. (2011) Mid1, a mechanosensitive calcium ion channel, affects growth, development, and ascospore discharge in the filamentous fungus Gibberella zeae. Eukaryot Cell 10: 832-841. doi: 10.1128/EC.00235-10
|
| [67] |
Hallen HE, Trail F (2008) The L-type calcium ion channel cch1 affects ascospore discharge and mycelial growth in the filamentous fungus Gibberella zeae (anamorph Fusarium graminearum). Eukaryot Cell 7: 415-424. doi: 10.1128/EC.00248-07
|
| [68] |
Bormann J, Tudzynski P (2009) Deletion of Mid1, a putative stretch-activated calcium channel in Claviceps purpurea, affects vegetative growth, cell wall synthesis and virulence. Microbiology 155: 3922-3933. doi: 10.1099/mic.0.030825-0
|
| [69] |
Zhou XL, Stumpf MA, Hoch HC, et al. (1991) A mechanosensitive channel in whole cells and in membrane patches of the fungus Uromyces. Science 253: 1415-1417. doi: 10.1126/science.1716786
|
| [70] |
Nakajima-Shimada J, Sakaguchi S, Tsuji F, et al. (2000) Ca2+ signal is generated only once in the mating pheromone response pathway in Saccharomyces cerevisiae. Cell Struct Funct 25: 125-131. doi: 10.1247/csf.25.125
|
| [71] |
Batiza A, Schulz T, Masson P (1996) Yeast respond to hypotonic shock with a calcium pulse. J Biol Chem 271: 23357-23362. doi: 10.1074/jbc.271.38.23357
|
| [72] |
Matsumoto T, Ellsmore A, Cessna S, et al. (2002) An osmotically induced cytosolic Ca2+ transient activates calcineurin signaling to mediate ion homeostasis and salt tolerance of Saccharomyces cerevisiae. J Biol Chem 277: 33075-33080. doi: 10.1074/jbc.M205037200
|
| [73] |
Peiter E, Fischer M, Sidaway K, et al. (2005) The Saccharomyces cerevisiae Ca2+ channel Cch1pMid1p is essential for tolerance to cold stress and iron toxicity. FEBS Lett 579: 5697-5703. doi: 10.1016/j.febslet.2005.09.058
|
| [74] |
Locke E, Bonilla M, Liang L, et al. (2000) A homolog of voltage-gated Ca2+ channels stimulated by depletion of secretory Ca2+ in yeast. Mol Cell Biol 20: 6686-6694. doi: 10.1128/MCB.20.18.6686-6694.2000
|
| [75] |
Rigamonti M, Groppi S, Belotti F, et al. (2015) Hypotonic stress-induced calcium signaling in Saccharomyces cerevisiae involves TRP-like transporters on the endoplasmic reticulum membrane. Cell Calcium 57: 57-68. doi: 10.1016/j.ceca.2014.12.003
|
| [76] | Courchesne WE, Vlasek C, Klukovich R, et al. (2011) Ethanol induces calcium influx via the Cch1-Mid1 transporter in Saccharomyces cerevisiae. Arch Microbiol 193: 323-334. |
| [77] |
Groppi S, Belotti F, Brand?o RL, et al. (2011) Glucose-induced calcium influx in budding yeast involves a novel calcium transport system and can activate calcineurin. Cell Calcium 49: 376-386. doi: 10.1016/j.ceca.2011.03.006
|
| [78] |
T?k��s-F��zesi M, Bedwell D, Repa I, et al. (2002) Hexose phosphorylation and the putative calcium channel component Mid1p are required for the hexose-induced transient elevation of cytosolic calcium response in Saccharomyces cerevisiae. Mol Microbiol 44: 1299-1308. doi: 10.1046/j.1365-2958.2002.02956.x
|
| [79] |
Tisi R, Baldassa S, Belotti F, et al. (2002) Phospholipase C is required for glucose-induced calcium influx in budding yeast. FEBS Lett 520: 133-138. doi: 10.1016/S0014-5793(02)02806-5
|
| [80] |
Viladevall L, Serrano R, Ruiz A, et al. (2004) Characterization of the calcium-mediated response to alkaline stress in Saccharomyces cerevisiae. J Biol Chem 279: 43614-43624. doi: 10.1074/jbc.M403606200
|
| [81] | Courchesne W, Ozturk S (2003) Amiodarone induces a caffeine-inhibited, MID1-depedent rise in free cytoplasmic calcium in Saccharomyces cerevisiae. Mol Microbiol 47: 223-234. |
| [82] | Muller E, Locke E, Cunningham K (2001) Differential regulation of two Ca2+ influx systems by pheromone signaling in Saccharomyces cerevisiae. Genetics 159: 1527-1538. |
| [83] |
Bonilla M, Cunningham K (2003) Mitogen-activated protein kinase stimulation of Ca2+ signaling is required for survival of endoplasmic reticulum stress in yeast. Mol Biol Cell 14: 4296-4305. doi: 10.1091/mbc.E03-02-0113
|
| [84] |
Jiang H, Shen Y, Liu W, et al. (2014) Deletion of the putative stretch-activated ion channel Mid1 is hypervirulent in Aspergillus fumigatus. Fungal Genet Biol 62: 62-70. doi: 10.1016/j.fgb.2013.11.003
|
| [85] |
Paidhungat M, Garrett S (1997) A homolog of mammalian, voltage-gated calcium channels mediates yeast pheromone-stimulated Ca2+ uptake and exacerbates the cdc1(Ts) growth defect. Mol Cell Biol 17: 6339-6347. doi: 10.1128/MCB.17.11.6339
|
| [86] |
Hong MP, Vu K, Bautos JM, et al. (2013) Activity of the calcium channel pore Cch1 is dependent on a modulatory region of the subunit Mid1 in Cryptococcus neoformans. Eukaryot Cell 12: 142-150. doi: 10.1128/EC.00130-12
|
| [87] |
Wang H, Liang Y, Zhang B, et al. (2011) Alkaline stress triggers an immediate calcium fluctuation in Candida albicans mediated by Rim101p and Crz1p transcription factors. FEMS Yeast Res 11: 430-439. doi: 10.1111/j.1567-1364.2011.00730.x
|
| [88] | Eilam Y, Chernichovsky D (1987) Uptake of Ca2+ driven by the membrane potential in energy-depleted yeast cells. J Gen Microbiol 133: 1641-1649. |
| [89] |
Eilam Y, Othman M (1990) Activation of Ca2+ influx by metabolic substrates in Saccharomyces cerevisiae: role of membrane potential and cellular ATP levels. J Gen Microbiol 136: 861-866. doi: 10.1099/00221287-136-5-861
|
| [90] |
Pereira M, Tisi R, Fietto L, et al. (2008) Carbonyl cyanide m-chlorophenylhydrazone induced calcium signaling and activation of plasma membrane H+-ATPase in the yeast Saccharomyces cerevisiae. FEMS Yeast Res 8: 622-630. doi: 10.1111/j.1567-1364.2008.00380.x
|
| [91] |
Stefan CP, Zhang N, Sokabe T, et al. (2013) Activation of an essential calcium signaling pathway in Saccharomyces cerevisiae by Kch1 and Kch2, putative low-affinity potassium transporters. Eukaryot Cell 12: 204-214. doi: 10.1128/EC.00299-12
|
| [92] |
Stefan CP, Cunningham KW (2013) Kch1 family proteins mediate essential responses to endoplasmic reticulum stresses in the yeasts Saccharomyces cerevisiae and Candida albicans. J Biol Chem 288: 34861-34870. doi: 10.1074/jbc.M113.508705
|
| [93] |
Maruoka T, Nagasoe Y, Inoue S, et al. (2002) Essential hydrophilic carboxyl-terminal regions including cysteine residues of the yeast stretch-activated calcium-permeable channel Mid1. J Biol Chem 277: 11645-11652. doi: 10.1074/jbc.M111603200
|
| [94] |
Possani LD, Becerril B, Delepierre M, et al. (1999) Scorpion toxins specific for Na+-channels. Eur J Biochem 264: 287-300. doi: 10.1046/j.1432-1327.1999.00625.x
|
| [95] |
Senatore A, Monteil A, van Minnen J, et al. (2013) NALCN ion channels have alternative selectivity filters resembling calcium channels or sodium channels. PLoS One 8: e55088. doi: 10.1371/journal.pone.0055088
|
| [96] | Ghezzi A, Liebeskind BJ, Thompson A, et al. (2014) Ancient association between cation leak channels and Mid1 proteins is conserved in fungi and animals. Front Mol Neurosci 7: 15. |
| [97] |
Teng J, Goto R, Iida K, et al. (2008) Ion-channel blocker sensitivity of voltage-gated calcium-channel homologue Cch1 in Saccharomyces cerevisiae. Microbiology 154: 3775-3781. doi: 10.1099/mic.0.2008/021089-0
|
| [98] |
Kmetzsch L, Staats CC, Rodrigues ML, et al. (2011) Calcium signaling components in the human pathogen: Cryptococcus neoformans. Commun Integr Biol 4: 186-187. doi: 10.4161/cib.4.2.14271
|
| [99] |
de Castro PA, Chiaratto J, Winkelstroter LK, et al. (2014) The involvement of the Mid1/Cch1/Yvc1 calcium channels in Aspergillus fumigatus virulence. PLoS One 9: e103957. doi: 10.1371/journal.pone.0103957
|
| [100] |
Yang M, Brand A, Srikantha T, et al. (2011) Fig1 Facilitates Calcium Influx and Localizes to Membranes Destined To Undergo Fusion during Mating in Candida albicans. Eukaryot Cell 10: 435-444. doi: 10.1128/EC.00145-10
|
| [101] |
Cavinder B, Trail F (2012) Role of Fig1, a component of the low-affinity calcium uptake system, in growth and sexual development of filamentous fungi. Eukaryot Cell 11: 978-988. doi: 10.1128/EC.00007-12
|
| [102] |
Muller E, Mackin N, Erdman S, et al. (2003) Fig1p facilitates Ca2+ influx and cell fusion during mating of Saccharomyces cerevisiae. J Biol Chem 278: 38461-38469. doi: 10.1074/jbc.M304089200
|
| [103] |
Zhang S, Zheng H, Long N, et al. (2014) FigA, a putative homolog of low-affinity calcium system member Fig1 in Saccharomyces cerevisiae, is involved in growth and asexual and sexual development in Aspergillus nidulans. Eukaryot Cell 13: 295-303. doi: 10.1128/EC.00257-13
|
| [104] |
Van Itallie CM, Anderson JM (2006) Claudins and epithelial paracellular transport. Annu Rev Physiol 68: 403-429. doi: 10.1146/annurev.physiol.68.040104.131404
|
| [105] | Gunzel D, Fromm M (2012) Claudins and other tight junction proteins. Compr Physiol 2: 1819-1852. |
| [106] |
Zhang N, Dudgeon D, Paliwal S, et al. (2006) Multiple signaling pathways regulate yeast cell death during the response to mating pheromones. Mol Biol Cell 17: 3409-3422. doi: 10.1091/mbc.E06-03-0177
|
| [107] | Brand A, Shanks S, Duncan VM, et al. (2007) Hyphal orientation of Candida albicans is regulated by a calcium-dependent mechanism. Curr Biol 17: 347-352. |
| [108] |
Venkatachalam K, Montell C (2007) TRP channels. Annu Rev Biochem 76: 387-417. doi: 10.1146/annurev.biochem.75.103004.142819
|
| [109] |
Vrenken KS, Jalink K, van Leeuwen FN, et al. (2016) Beyond ion-conduction: Channel-dependent and -independent roles of TRP channels during development and tissue homeostasis. Biochim Biophys Acta 1863: 1436-1446. doi: 10.1016/j.bbamcr.2015.11.008
|
| [110] |
Palmer C, Zhou X, Lin J, et al. (2001) A TRP homolog in Saccharomyces cerevisiae forms an intracellular Ca2+-permeable channel in the yeast vacuolar membrane. Proc Natl Acad Sci U S A 98: 7801-7805. doi: 10.1073/pnas.141036198
|
| [111] |
Denis V, Cyert M (2002) Internal Ca2+ release in yeast is triggered by hypertonic shock and mediated by a TRP channel homologue. J Cell Biol 156: 29-34. doi: 10.1083/jcb.200111004
|
| [112] |
Zhou X, Batiza A, Loukin S, et al. (2003) The transient receptor potential channel on the yeast vacuole is mechanosensitive. Proc Natl Acad Sci U S A 100: 7105-7110. doi: 10.1073/pnas.1230540100
|
| [113] |
Chang Y, Schlenstedt G, Flockerzi V, et al. (2010) Properties of the intracellular transient receptor potential (TRP) channel in yeast, Yvc1. FEBS Lett 584: 2028-2032. doi: 10.1016/j.febslet.2009.12.035
|
| [114] |
Su Z, Zhou X, Loukin S, et al. (2009) Mechanical force and cytoplasmic Ca2+ activate yeast TRPY1 in parallel. J Membr Biol 227: 141-150. doi: 10.1007/s00232-009-9153-9
|
| [115] |
Su Z, Anishkin A, Kung C, et al. (2011) The core domain as the force sensor of the yeast mechanosensitive TRP channel. J Gen Physiol 138: 627-640. doi: 10.1085/jgp.201110693
|
| [116] |
Bouillet LE, Cardoso AS, Perovano E, et al. (2012) The involvement of calcium carriers and of the vacuole in the glucose-induced calcium signaling and activation of the plasma membrane H+-ATPase in Saccharomyces cerevisiae cells. Cell Calcium 51: 72-81. doi: 10.1016/j.ceca.2011.10.008
|
| [117] |
Yu Q, Wang F, Zhao Q, et al. (2014) A novel role of the vacuolar calcium channel Yvc1 in stress response, morphogenesis and pathogenicity of Candida albicans. Int J Med Microbiol 304: 339-350. doi: 10.1016/j.ijmm.2013.11.022
|
| [118] |
Prole DL, Taylor CW (2012) Identification and analysis of cation channel homologues in human pathogenic fungi. PLoS One 7: e42404. doi: 10.1371/journal.pone.0042404
|
| [119] |
Boratyn GM, Camacho C, Cooper PS, et al. (2013) BLAST: a more efficient report with usability improvements. Nucleic Acids Res 41: W29-33. doi: 10.1093/nar/gkt282
|
| [120] |
Goncalves AP, Cordeiro JM, Monteiro J, et al. (2014) Activation of a TRP-like channel and intracellular Ca2+ dynamics during phospholipase-C-mediated cell death. J Cell Sci 127: 3817-3829. doi: 10.1242/jcs.152058
|
| [121] |
Bonilla M, Nastase K, Cunningham K (2002) Essential role of calcineurin in response to endoplasmic reticulum stress. EMBO J 21: 2343-2353. doi: 10.1093/emboj/21.10.2343
|
| [122] |
Hong MP, Vu K, Bautos J, et al. (2010) Cch1 restores intracellular Ca2+ in fungal cells during endoplasmic reticulum stress. J Biol Chem 285: 10951-10958. doi: 10.1074/jbc.M109.056218
|
| [123] |
Okada Y, Maeno E, Shimizu T, et al. (2001) Receptor-mediated control of regulatory volume decrease (RVD) and apoptotic volume decrease (AVD). J Physiol 532: 3-16. doi: 10.1111/j.1469-7793.2001.0003g.x
|
| [124] |
Becker D, Blase C, Bereiter-Hahn J, et al. (2005) TRPV4 exhibits a functional role in cell-volume regulation. J Cell Sci 118: 2435-2440. doi: 10.1242/jcs.02372
|
| [125] | Numata T, Shimizu T, Okada Y (2007) TRPM7 is a stretch- and swelling-activated cation channel involved in volume regulation in human epithelial cells. Am J Physiol Cell Physiol 292: C460-467. |
| [126] |
Berrier C, Besnard M, Ajouz B, et al. (1996) Multiple mechanosensitive ion channels from Escherichia coli, activated at different thresholds of applied pressure. J Membr Biol 151: 175-187. doi: 10.1007/s002329900068
|
| [127] |
Rigamonti M, Belotti F, Martegani E, et al. (2013) Novel role for known and unknown calcium transport involved proteins in Saccharomyces cerevisiae. Yeast 30: 157. doi: 10.1002/yea.2946
|
| [128] |
Protchenko O, Rodriguez-Suarez R, Androphy R, et al. (2006) A screen for genes of heme uptake identifies the FLC family required for import of FAD into the endoplasmic reticulum. J Biol Chem 281: 21445-21457. doi: 10.1074/jbc.M512812200
|
| [129] |
Palmer CP, Aydar E, Djamgoz MB (2005) A microbial TRP-like polycystic-kidney-disease-related ion channel gene. Biochem J 387: 211-219. doi: 10.1042/BJ20041710
|
| [130] |
Aydar E, Palmer CP (2009) Polycystic kidney disease channel and synaptotagmin homologues play roles in Schizosaccharomyces pombe cell wall synthesis/repair and membrane protein trafficking. J Membr Biol 229: 141-152. doi: 10.1007/s00232-009-9180-6
|
| [131] | Rzhetsky A, Nei M (1994) METREE: a program package for inferring and testing minimum-evolution trees. Comput Appl Biosci 10: 409-412. |
| [132] |
Tamura K, Stecher G, Peterson D, et al. (2013) MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol 30: 2725-2729. doi: 10.1093/molbev/mst197
|
| [133] |
Efron B, Halloran E, Holmes S (1996) Bootstrap confidence levels for phylogenetic trees. Proc Natl Acad Sci U S A 93: 13429-13434. doi: 10.1073/pnas.93.23.13429
|
| [134] | Jones DT, Taylor WR, Thornton JM (1992) The rapid generation of mutation data matrices from protein sequences. Comput Appl Biosci 8: 275-282. |
| [135] | Nei M, Kumar S (2000) Molecular Evolution and Phylogenetics, Oxford University Press, New York. |
| [136] | Saitou N, Nei M (1987) The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 4: 406-425. |
| [137] |
Hsiang T, Baillie DL (2005) Comparison of the yeast proteome to other fungal genomes to find core fungal genes. J Mol Evol 60: 475-483. doi: 10.1007/s00239-004-0218-1
|
| [138] |
Bok JW, Sone T, Silverman-Gavrila LB, et al. (2001) Structure and function analysis of the calcium-related gene spray in Neurospora crassa. Fungal Genet Biol 32: 145-158. doi: 10.1006/fgbi.2000.1259
|
| [139] |
Stephenson KS, Gow NA, Davidson FA, et al. (2014) Regulation of vectorial supply of vesicles to the hyphal tip determines thigmotropism in Neurospora crassa. Fungal Biol 118: 287-294. doi: 10.1016/j.funbio.2013.12.007
|
| [140] |
Colletti GA, Kiselyov K (2011) TRPML1. Adv Exp Med Biol 704: 209-219. doi: 10.1007/978-94-007-0265-3_11
|
| [141] |
Puertollano R, Kiselyov K (2009) TRPMLs: in sickness and in health. Am J Physiol Renal Physiol 296: F1245-1254. doi: 10.1152/ajprenal.90522.2008
|
| [142] | Fares H, Greenwald I (2001) Regulation of endocytosis by CUP-5, the Caenorhabditis elegans mucolipin-1 homolog. Nat Genet 28: 64-68. |
| [143] |
Lima WC, Leuba F, Soldati T, et al. (2012) Mucolipin controls lysosome exocytosis in Dictyostelium. J Cell Sci 125: 2315-2322. doi: 10.1242/jcs.100362
|
| [144] | Dong XP, Shen D, Wang X, et al. (2010) PI(3,5)P(2) controls membrane trafficking by direct activation of mucolipin Ca2+ release channels in the endolysosome. Nat Commun 1: 38. |
| [145] |
Ma Y, Sugiura R, Koike A, et al. (2011) Transient receptor potential (TRP) and Cch1-Yam8 channels play key roles in the regulation of cytoplasmic Ca2+ in fission yeast. PLoS One 6: e22421. doi: 10.1371/journal.pone.0022421
|
| [146] | Bonora M, Pinton P (2014) The mitochondrial permeability transition pore and cancer: molecular mechanisms involved in cell death. Front Oncol 4: 302. |
| [147] |
Carraro M, Giorgio V, Sileikyte J, et al. (2014) Channel formation by yeast F-ATP synthase and the role of dimerization in the mitochondrial permeability transition. J Biol Chem 289: 15980-15985. doi: 10.1074/jbc.C114.559633
|
| [148] |
De Stefani D, Raffaello A, Teardo E, et al. (2011) A forty-kilodalton protein of the inner membrane is the mitochondrial calcium uniporter. Nature 476: 336-340. doi: 10.1038/nature10230
|
| [149] |
Baughman JM, Perocchi F, Girgis HS, et al. (2011) Integrative genomics identifies MCU as an essential component of the mitochondrial calcium uniporter. Nature 476: 341-345. doi: 10.1038/nature10234
|
| [150] |
Marchi S, Pinton P (2014) The mitochondrial calcium uniporter complex: molecular components, structure and physiopathological implications. J Physiol 592: 829-839. doi: 10.1113/jphysiol.2013.268235
|
| [151] |
Uribe S, Rangel P, Pardo J (1992) Interactions of calcium with yeast mitochondria. Cell Calcium 13: 211-217. doi: 10.1016/0143-4160(92)90009-H
|
| [152] |
Nakayama Y, Yoshimura K, Iida H (2012) Organellar mechanosensitive channels in fission yeast regulate the hypo-osmotic shock response. Nat Commun 3: 1020. doi: 10.1038/ncomms2014
|
| [153] |
Nakayama Y, Hirata A, Iida H (2014) Mechanosensitive channels Msy1 and Msy2 are required for maintaining organelle integrity upon hypoosmotic shock in Schizosaccharomyces pombe. FEMS Yeast Res 14: 992-994. doi: 10.1111/1567-1364.12181
|
| [154] |
Gustin MC, Zhou XL, Martinac B, et al. (1988) A mechanosensitive ion channel in the yeast plasma membrane. Science 242: 762-765. doi: 10.1126/science.2460920
|
| [155] |
Triggle DJ (2006) L-type calcium channels. Curr Pharm Des 12: 443-457. doi: 10.2174/138161206775474503
|
| [156] |
Jiang L, Alber J, Wang J, et al. (2012) The Candida albicans plasma membrane protein Rch1p, a member of the vertebrate SLC10 carrier family, is a novel regulator of cytosolic Ca2+ homoeostasis. Biochem J 444: 497-502. doi: 10.1042/BJ20112166
|
| [157] |
Zhao Y, Yan H, Happeck R, et al. (2016) The plasma membrane protein Rch1 is a negative regulator of cytosolic calcium homeostasis and positively regulated by the calcium/calcineurin signaling pathway in budding yeast. Eur J Cell Biol 95: 164-174. doi: 10.1016/j.ejcb.2016.01.001
|
| [158] |
Kraus P, Heitman J (2003) Coping with stress: calmodulin and calcineurin in model and pathogenic fungi. Biochem Biophys Res Commun 311: 1151-1157. doi: 10.1016/S0006-291X(03)01528-6
|
| [159] |
Lewit-Bentley A, Rety S (2000) EF-hand calcium-binding proteins. Curr Opin Struct Biol 10: 637-643. doi: 10.1016/S0959-440X(00)00142-1
|
| [160] | Matsuura I, Ishihara K, Nakai Y, et al. (1991) A site-directed mutagenesis study of yeast calmodulin. J Biochem 109: 190-197. |
| [161] |
Cyert M (2001) Genetic analysis of calmodulin and its targets in Saccharomyces cerevisiae. Annu Rev Genet 35: 647-672. doi: 10.1146/annurev.genet.35.102401.091302
|
| [162] |
Cruz MC, Goldstein AL, Blankenship JR, et al. (2002) Calcineurin is essential for survival during membrane stress in Candida albicans. EMBO J 21: 546-559. doi: 10.1093/emboj/21.4.546
|
| [163] |
Kontoyiannis DP, Lewis RE, Osherov N, et al. (2003) Combination of caspofungin with inhibitors of the calcineurin pathway attenuates growth in vitro in Aspergillus species. J Antimicrob Chemother 51: 313-316. doi: 10.1093/jac/dkg090
|
| [164] |
Steinbach WJ, Reedy JL, Cramer RA Jr, et al. (2007) Harnessing calcineurin as a novel anti-infective agent against invasive fungal infections. Nat Rev Microbiol 5: 418-430. doi: 10.1038/nrmicro1680
|
| [165] |
Rispail N, Soanes DM, Ant C, et al. (2009) Comparative genomics of MAP kinase and calcium-calcineurin signalling components in plant and human pathogenic fungi. Fungal Genet Biol 46: 287-298. doi: 10.1016/j.fgb.2009.01.002
|
| [166] |
Moser M, Lee S, Klevit R, et al. (1995) Ca2+ binding to calmodulin and its role in Schizosaccharomyces pombe as revealed by mutagenesis and NMR spectroscopy. J Biol Chem 270: 20643-20652. doi: 10.1074/jbc.270.35.20643
|
| [167] |
Joseph J, Means A (2002) Calcium binding is required for calmodulin function in Aspergillus nidulans. Eukaryot Cell 1: 119-125. doi: 10.1128/EC.01.1.119-125.2002
|
| [168] |
Davis T, Urdea M, Masiarz F, et al. (1986) Isolation of the yeast calmodulin gene: calmodulin is an essential protein. Cell 47: 423-431. doi: 10.1016/0092-8674(86)90599-4
|
| [169] |
Geiser J, van Tuinen D, Brockerhoff S, et al. (1991) Can calmodulin function without binding calcium? Cell 65: 949-959. doi: 10.1016/0092-8674(91)90547-C
|
| [170] |
Ohya Y, Botstein D (1994) Diverse essential functions revealed by complementing yeast calmodulin mutants. Science 263: 963-966. doi: 10.1126/science.8310294
|
| [171] |
Uttenweiler A, Schwarz H, Mayer A (2005) Microautophagic vacuole invagination requires calmodulin in a Ca2+-independent function. J Biol Chem 280: 33289-33297. doi: 10.1074/jbc.M506086200
|
| [172] |
Geiser JR, Sundberg HA, Chang BH, et al. (1993) The essential mitotic target of calmodulin is the 110-kilodalton component of the spindle pole body in Saccharomyces cerevisiae. Mol Cell Biol 13: 7913-7924. doi: 10.1128/MCB.13.12.7913
|
| [173] |
Brockerhoff S, Stevens R, Davis T (1994) The unconventional myosin, Myo2p, is a calmodulin target at sites of cell growth in Saccharomyces cerevisiae. J Cell Biol 124: 315-323. doi: 10.1083/jcb.124.3.315
|
| [174] |
de Carvalho MJ, Amorim Jesuino RS, Daher BS, et al. (2003) Functional and genetic characterization of calmodulin from the dimorphic and pathogenic fungus Paracoccidioides brasiliensis. Fungal Genet Biol 39: 204-210. doi: 10.1016/S1087-1845(03)00044-6
|
| [175] | Lee SC, Lee YH (1998) Calcium/calmodulin-dependent signaling for appressorium formation in the plant pathogenic fungus Magnaporthe grisea. Mol Cells 8: 698-704. |
| [176] |
Warwar V, Oved S, Dickman MB (2000) Antisense expression of the calmodulin gene from Colletotrichum trifolii impairs prepenetration development(1). FEMS Microbiol Lett 191: 213-219. doi: 10.1111/j.1574-6968.2000.tb09342.x
|
| [177] |
Kraus PR, Nichols CB, Heitman J (2005) Calcium- and calcineurin-independent roles for calmodulin in Cryptococcus neoformans morphogenesis and high-temperature growth. Eukaryot Cell 4: 1079-1087. doi: 10.1128/EC.4.6.1079-1087.2005
|
| [178] | Ohya Y, Kawasaki H, Suzuki K, et al. (1991) Two yeast genes encoding calmodulin-dependent protein kinases. Isolation, sequencing and bacterial expressions of CMK1 and CMK2. J Biol Chem 266: 12784-12794. |
| [179] |
Cyert M, Kunisawa R, Kaim D, et al. (1991) Yeast has homologs (CNA1 and CNA2 gene products) of mammalian calcineurin, a calmodulin-regulated phosphoprotein phosphatase. Proc Natl Acad Sci U S A 88: 7376-7380. doi: 10.1073/pnas.88.16.7376
|
| [180] |
Bodvard K, Jorhov A, Blomberg A, et al. (2013) The yeast transcription factor Crz1 is activated by light in a Ca2+/calcineurin-dependent and PKA-independent manner. PLoS One 8: e53404. doi: 10.1371/journal.pone.0053404
|
| [181] |
Farcasanu IC, Mitrica R, Cristache L, et al. (2013) Optical manipulation of Saccharomyces cerevisiae cells reveals that green light protection against UV irradiation is favored by low Ca2+ and requires intact UPR pathway. FEBS Lett 587: 3514-3521. doi: 10.1016/j.febslet.2013.09.008
|
| [182] |
Hirata D, Harada S, Namba H, et al. (1995) Adaptation to high-salt stress in Saccharomyces cerevisiae is regulated by Ca2+/calmodulin-dependent phosphoprotein phosphatase (calcineurin) and cAMP-dependent protein kinase. Mol Gen Genet 249: 257-264. doi: 10.1007/BF00290525
|
| [183] |
Iida H, Ohya Y, Anraku Y (1995) Calmodulin-dependent protein kinase II and calmodulin are required for induced thermotolerance in Saccharomyces cerevisiae. Curr Genet 27: 190-193. doi: 10.1007/BF00313434
|
| [184] |
Moser M, Geiser J, Davis T (1996) Ca2+-calmodulin promotes survival of pheromone-induced growth arrest by activation of calcineurin and Ca2+-calmodulin-dependent protein kinase. Mol Cell Biol 16: 4824-4831. doi: 10.1128/MCB.16.9.4824
|
| [185] |
Breuder T, Hemenway CS, Movva NR, et al. (1994) Calcineurin is essential in cyclosporin A- and FK506-sensitive yeast strains. Proc Natl Acad Sci U S A 91: 5372-5376. doi: 10.1073/pnas.91.12.5372
|
| [186] | Hemenway CS, Dolinski K, Cardenas ME, et al. (1995) vph6 mutants of Saccharomyces cerevisiae require calcineurin for growth and are defective in vacuolar H+-ATPase assembly. Genetics 141: 833-844. |
| [187] | Holyoak CD, Thompson S, Ortiz Calderon C, et al. (2000) Loss of Cmk1 Ca2+-calmodulin-dependent protein kinase in yeast results in constitutive weak organic acid resistance, associated with a post-transcriptional activation of the Pdr12 ATP-binding cassette transporter. Mol Microbiol 37: 595-605. |
| [188] |
Mendoza I, Quintero FJ, Bressan RA, et al. (1996) Activated calcineurin confers high tolerance to ion stress and alters the budding pattern and cell morphology of yeast cells. J Biol Chem 271: 23061-23067. doi: 10.1074/jbc.271.38.23061
|
| [189] |
Cyert M (2003) Calcineurin signaling in Saccharomyces cerevisiae: how yeast go crazy in response to stress. Biochem Biophys Res Commun 311: 1143-1150. doi: 10.1016/S0006-291X(03)01552-3
|
| [190] |
Stathopoulos-Gerontides A, Guo J, Cyert M (1999) Yeast calcineurin regulates nuclear localization of the Crz1p transcription factor through dephosphorylation. Genes Dev 13: 798-803. doi: 10.1101/gad.13.7.798
|
| [191] |
Boustany L, Cyert M (2002) Calcineurin-dependent regulation of Crz1p nuclear export requires Msn5p and a conserved calcineurin docking site. Genes Dev 16: 608-619. doi: 10.1101/gad.967602
|
| [192] |
Yoshimoto H, Saltsman K, Gasch A, et al. (2002) Genome-wide analysis of gene expression regulated by the calcineurin/Crz1p signaling pathway in Saccharomyces cerevisiae. J Biol Chem 277: 31079-31088. doi: 10.1074/jbc.M202718200
|
| [193] |
Marchi V, Sorin A, Wei Y, et al. (1999) Induction of vacuolar Ca2+-ATPase and H+/Ca2+ exchange activity in yeast mutants lacking Pmr1, the Golgi Ca2+-ATPase. FEBS Lett 454: 181-186. doi: 10.1016/S0014-5793(99)00803-0
|
| [194] |
Cunningham K, Fink G (1996) Calcineurin inhibits VCX1-dependent H+/Ca2+ exchange and induces Ca2+ ATPases in Saccharomyces cerevisiae. Mol Cell Biol 16: 2226-2237. doi: 10.1128/MCB.16.5.2226
|
| [195] |
Zhao C, Jung U, Garrett-Engele P, et al. (1998) Temperature-induced expression of yeast FKS2 is under the dual control of protein kinase C and calcineurin. Mol Cell Biol 18: 1013-1022. doi: 10.1128/MCB.18.2.1013
|
| [196] |
Hernandez-Lopez M. J, Panadero J, Prieto JA, et al. (2006) Regulation of salt tolerance by Torulaspora delbrueckii calcineurin target Crz1p. Eukaryot Cell 5: 469-479. doi: 10.1128/EC.5.3.469-479.2006
|
| [197] |
Chen YL, Brand A, Morrison EL, et al. (2011) Calcineurin controls drug tolerance, hyphal growth, and virulence in Candida dubliniensis. Eukaryot Cell 10: 803-819. doi: 10.1128/EC.00310-10
|
| [198] |
Chen YL, Yu SJ, Huang HY, et al. (2014) Calcineurin controls hyphal growth, virulence, and drug tolerance of Candida tropicalis. Eukaryot Cell 13: 844-854. doi: 10.1128/EC.00302-13
|
| [199] | Juvvadi PR, Kuroki Y, Arioka M, et al. (2003) Functional analysis of the calcineurin-encoding gene cnaA from Aspergillus oryzae: evidence for its putative role in stress adaptation. Arch Microbiol 179: 416-422. |
| [200] |
Kothe GO, Free SJ (1998) Calcineurin subunit B is required for normal vegetative growth in Neurospora crassa. Fungal Genet Biol 23: 248-258. doi: 10.1006/fgbi.1998.1037
|
| [201] |
Harel A, Bercovich S, Yarden O (2006) Calcineurin is required for sclerotial development and pathogenicity of Sclerotinia sclerotiorum in an oxalic acid-independent manner. Mol Plant Microbe Interact 19: 682-693. doi: 10.1094/MPMI-19-0682
|
| [202] | Choi J, Kim Y, Lee Y (2009) Functional analysis of MCNA, a gene encoding a catalytic subunit of calcineurin, in the rice blast fungus Magnaporthe oryzae. J Microbiol Biotechnol 19: 11-16. |
| [203] |
Cramer RA Jr, Perfect BZ, Pinchai N, et al. (2008) Calcineurin target CrzA regulates conidial germination, hyphal growth, and pathogenesis of Aspergillus fumigatus. Eukaryot Cell 7: 1085-1097. doi: 10.1128/EC.00086-08
|
| [204] |
Steinbach WJ, Cramer RA Jr, Perfect BZ et al. (2006) Calcineurin controls growth, morphology, and pathogenicity in Aspergillus fumigatus. Eukaryot Cell 5: 1091-1103. doi: 10.1128/EC.00139-06
|
| [205] |
Juvvadi PR, Fortwendel JR, Pinchai N, et al. (2008) Calcineurin localizes to the hyphal septum in Aspergillus fumigatus: implications for septum formation and conidiophore development. Eukaryot Cell 7: 1606-1610. doi: 10.1128/EC.00200-08
|
| [206] |
Juvvadi PR, Fortwendel JR, Rogg LE, et al. (2011) Localization and activity of the calcineurin catalytic and regulatory subunit complex at the septum is essential for hyphal elongation and proper septation in Aspergillus fumigatus. Mol Microbiol 82: 1235-1259. doi: 10.1111/j.1365-2958.2011.07886.x
|
| [207] |
Miyakawa T, Mizunuma M (2007) Physiological roles of calcineurin in Saccharomyces cerevisiae with special emphasis on its roles in G2/M cell-cycle regulation. Biosci Biotechnol Biochem 71: 633-645. doi: 10.1271/bbb.60495
|
| [208] |
Wu H, Peisley A, Graef IA, et al. (2007) NFAT signaling and the invention of vertebrates. Trends Cell Biol 17: 251-260. doi: 10.1016/j.tcb.2007.04.006
|
| [209] | Hirayama S, Sugiura R, Lu Y, et al. (2003) Zinc finger protein Prz1 regulates Ca2+ but not Cl- homeostasis in fission yeast. Identification of distinct branches of calcineurin signaling pathway in fission yeast. J Biol Chem 278: 18078-18084. |
| [210] |
Soriani FM, Malavazi I, da Silva Ferreira ME, et al. (2008) Functional characterization of the Aspergillus fumigatus CRZ1 homologue, CrzA. Mol Microbiol 67: 1274-1291. doi: 10.1111/j.1365-2958.2008.06122.x
|
| [211] |
Thewes S (2014) Calcineurin-Crz1 signaling in lower eukaryotes. Eukaryot Cell 13: 694-705. doi: 10.1128/EC.00038-14
|
| [212] |
Zhang H, Zhao Q, Liu K, et al. (2009) MgCRZ1, a transcription factor of Magnaporthe grisea, controls growth, development and is involved in full virulence. FEMS Microbiol Lett 293: 160-169. doi: 10.1111/j.1574-6968.2009.01524.x
|
| [213] |
Choi J, Kim Y, Kim S, et al. (2009) MoCRZ1, a gene encoding a calcineurin-responsive transcription factor, regulates fungal growth and pathogenicity of Magnaporthe oryzae. Fungal Genet Biol 46: 243-254. doi: 10.1016/j.fgb.2008.11.010
|
| [214] |
Adler A, Park YD, Larsen P, et al. (2011) A novel specificity protein 1 (SP1)-like gene regulating protein kinase C-1 (Pkc1)-dependent cell wall integrity and virulence factors in Cryptococcus neoformans. J Biol Chem 286: 20977-20990. doi: 10.1074/jbc.M111.230268
|
| [215] |
Lev S, Desmarini D, Chayakulkeeree M, et al. (2012) The Crz1/Sp1 transcription factor of Cryptococcus neoformans is activated by calcineurin and regulates cell wall integrity. PLoS One 7: e51403. doi: 10.1371/journal.pone.0051403
|
| [216] | Moranova Z, Virtudazo E, Hricova K, et al. (2014) The CRZ1/SP1-like gene links survival under limited aeration, cell integrity and biofilm formation in the pathogenic yeast Cryptococcus neoformans. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub 158: 212-220. |
| [217] |
Santos M, de Larrinoa I (2005) Functional characterization of the Candida albicans CRZ1 gene encoding a calcineurin-regulated transcription factor. Curr Genet 48: 88-100. doi: 10.1007/s00294-005-0003-8
|
| [218] |
Karababa M, Valentino E, Pardini G, et al. (2006) CRZ1, a target of the calcineurin pathway in Candida albicans. Mol Microbiol 59: 1429-1451. doi: 10.1111/j.1365-2958.2005.05037.x
|
| [219] |
Chen YL, Konieczka JH, Springer DJ, et al. (2012) Convergent Evolution of Calcineurin Pathway Roles in Thermotolerance and Virulence in Candida glabrata. G3 (Bethesda) 2: 675-691. doi: 10.1534/g3.112.002279
|
| [220] |
Zhang J, Silao FG, Bigol UG, et al. (2012) Calcineurin is required for pseudohyphal growth, virulence, and drug resistance in Candida lusitaniae. PLoS One 7: e44192. doi: 10.1371/journal.pone.0044192
|
| [221] |
Araki Y, Wu H, Kitagaki H, et al. (2009) Ethanol stress stimulates the Ca2+-mediated calcineurin/Crz1 pathway in Saccharomyces cerevisiae. J Biosci Bioeng 107: 1-6. doi: 10.1016/j.jbiosc.2008.09.005
|
| [222] |
Zhang T, Xu Q, Sun X, et al. (2013) The calcineurin-responsive transcription factor Crz1 is required for conidation, full virulence and DMI resistance in Penicillium digitatum. Microbiol Res 168: 211-222. doi: 10.1016/j.micres.2012.11.006
|
| [223] |
Calvo IA, Gabrielli N, Iglesias-Baena I, et al. (2009) Genome-wide screen of genes required for caffeine tolerance in fission yeast. PLoS One 4: e6619. doi: 10.1371/journal.pone.0006619
|
| [224] |
Kafadar K, Cyert M (2004) Integration of stress responses: modulation of calcineurin signaling in Saccharomyces cerevisiae by protein kinase A. Eukaryot Cell 3: 1147-1153. doi: 10.1128/EC.3.5.1147-1153.2004
|
| [225] |
Kafadar K, Zhu H, Snyder M, et al. (2003) Negative regulation of calcineurin signaling by Hrr25p, a yeast homolog of casein kinase I. Genes Dev 17: 2698-2708. doi: 10.1101/gad.1140603
|
| [226] |
Koike A, Kato T, Sugiura R, et al. (2012) Genetic screening for regulators of Prz1, a transcriptional factor acting downstream of calcineurin in fission yeast. J Biol Chem 287: 19294-19303. doi: 10.1074/jbc.M111.310615
|
| [227] |
Williams K, Cyert M (2001) The eukaryotic response regulator Skn7p regulates calcineurin signaling through stabilization of Crz1p. EMBO J 20: 3473-3483. doi: 10.1093/emboj/20.13.3473
|
| [228] |
Hernandez-Ortiz P, Espeso EA (2013) Phospho-regulation and nucleocytoplasmic trafficking of CrzA in response to calcium and alkaline-pH stress in Aspergillus nidulans. Mol Microbiol 89: 532-551. doi: 10.1111/mmi.12294
|
| [229] |
da Silva Ferreira ME, Heinekamp T, Hartl A, et al. (2007) Functional characterization of the Aspergillus fumigatus calcineurin. Fungal Genet Biol 44: 219-230. doi: 10.1016/j.fgb.2006.08.004
|
| [230] |
Ruiz A, Serrano R, Ari?o J (2008) Direct regulation of genes involved in glucose utilization by the calcium/calcineurin pathway. J Biol Chem 283: 13923-13933. doi: 10.1074/jbc.M708683200
|
| [231] |
Rodriguez C, Galindo LR, Siverio JM (2013) Nitrogen-dependent calcineurin activation in the yeast Hansenula polymorpha. Fungal Genet Biol 53: 34-41. doi: 10.1016/j.fgb.2013.01.007
|
| [232] |
Li H, Rao A, Hogan PG (2011) Interaction of calcineurin with substrates and targeting proteins. Trends Cell Biol 21: 91-103. doi: 10.1016/j.tcb.2010.09.011
|
| [233] |
Gorlach J, Fox DS, Cutler NS, et al. (2000) Identification and characterization of a highly conserved calcineurin binding protein, CBP1/calcipressin in Cryptococcus neoformans. EMBO J 19: 3618-3629. doi: 10.1093/emboj/19.14.3618
|
| [234] |
Fuentes JJ, Pritchard MA, Estivill X (1997) Genomic organization, alternative splicing, and expression patterns of the DSCR1 (Down syndrome candidate region 1) gene. Genomics 44: 358-361. doi: 10.1006/geno.1997.4866
|
| [235] |
Hilioti Z, Gallagher DA, Low-Nam ST, et al. (2004) GSK-3 kinases enhance calcineurin signaling by phosphorylation of RCNs. Genes Dev 18: 35-47. doi: 10.1101/gad.1159204
|
| [236] |
Mehta S, Li H, Hogan P, et al. (2009) Domain architecture of the regulators of calcineurin (RCANs) and identification of a divergent RCAN in yeast. Mol Cell Biol 29: 2777-2793. doi: 10.1128/MCB.01197-08
|
| [237] |
Kishi T, Ikeda A, Nagao R, et al. (2007) The SCFCdc4 ubiquitin ligase regulates calcineurin signaling through degradation of phosphorylated Rcn1, an inhibitor of calcineurin. Proc Natl Acad Sci U S A 104: 17418-17423. doi: 10.1073/pnas.0704951104
|
| [238] | Rusnak F, Mertz P (2000) Calcineurin: form and function. Physiol Rev 80: 1483-1521. |
| [239] |
Connolly S, Kingsbury T (2012) Regulatory subunit myristoylation antagonizes calcineurin phosphatase activation in yeast. J Biol Chem 287: 39361-39368. doi: 10.1074/jbc.M112.366617
|
| [240] |
Miranda-Saavedra D, Barton GJ (2007) Classification and functional annotation of eukaryotic protein kinases. Proteins 68: 893-914. doi: 10.1002/prot.21444
|
| [241] | Pausch M, Kaim D, Kunisawa R, et al. (1991) Multiple Ca2+/calmodulin-dependent protein kinase genes in a unicellular eukaryote. EMBO J 10: 1511-1522. |
| [242] |
Dudgeon D, Zhang N, Ositelu O, et al. (2008) Nonapoptotic death of Saccharomyces cerevisiae cells that is stimulated by Hsp90 and inhibited by calcineurin and Cmk2 in response to endoplasmic reticulum stresses. Eukaryot Cell 7: 2037-2051. doi: 10.1128/EC.00291-08
|
| [243] |
Rasmussen CD (2000) Cloning of a calmodulin kinase I homologue from Schizosaccharomyces pombe. J Biol Chem 275: 685-690. doi: 10.1074/jbc.275.1.685
|
| [244] |
Sanchez-Piris M, Posas F, Alemany V, et al. (2002) The serine/threonine kinase Cmk2 is required for oxidative stress response in fission yeast. J Biol Chem 277: 17722-17727. doi: 10.1074/jbc.M200104200
|
| [245] |
Ding X, Yu Q, Zhang B, et al. (2014) The type II Ca2+/calmodulin-dependent protein kinases are involved in the regulation of cell wall integrity and oxidative stress response in Candida albicans. Biochem Biophys Res Commun 446: 1073-1078. doi: 10.1016/j.bbrc.2014.03.059
|
| [246] |
Yang Y, Cheng P, Zhi G, et al. (2001) Identification of a calcium/calmodulin-dependent protein kinase that phosphorylates the Neurospora circadian clock protein FREQUENCY. J Biol Chem 276: 41064-41072. doi: 10.1074/jbc.M106905200
|
| [247] |
Dayton JS, Means AR (1996) Ca2+/calmodulin-dependent kinase is essential for both growth and nuclear division in Aspergillus nidulans. Mol Biol Cell 7: 1511-1519. doi: 10.1091/mbc.7.10.1511
|
| [248] |
Liu XH, Lu JP, Dong B, et al. (2010) Disruption of MoCMK1, encoding a putative calcium/calmodulin-dependent kinase, in Magnaporthe oryzae. Microbiol Res 165: 402-410. doi: 10.1016/j.micres.2009.08.007
|
| [249] |
Hanyu Y, Imai KK, Kawasaki Y, et al. (2009) Schizosaccharomyces pombe cell division cycle under limited glucose requires Ssp1 kinase, the putative CaMKK, and Sds23, a PP2A-related phosphatase inhibitor. Genes Cells 14: 539-554. doi: 10.1111/j.1365-2443.2009.01290.x
|
| [250] |
Gomez-Hierro A, Lambea E, Gimenez-Zaragoza D, et al. (2015) Ssp1 CaMKK: A Sensor of Actin Polarization That Controls Mitotic Commitment through Srk1 in Schizosaccharomyces pombe. PLoS One 10: e0143037. doi: 10.1371/journal.pone.0143037
|
| [251] |
Cisneros-Barroso E, Yance-Chavez T, Kito A, et al. (2014) Negative feedback regulation of calcineurin-dependent Prz1 transcription factor by the CaMKK-CaMK1 axis in fission yeast. Nucleic Acids Res 42: 9573-9587. doi: 10.1093/nar/gku684
|
| [252] |
Yu Q, Wang H, Cheng X, et al. (2012) Roles of Cch1 and Mid1 in morphogenesis, oxidative stress response and virulence in Candida albicans. Mycopathologia 174: 359-369. doi: 10.1007/s11046-012-9569-0
|
| [253] |
Eilam Y, Othman M, Halachmi D (1990) Transient increase in Ca2+ influx in Saccharomyces cerevisiae in response to glucose: effects of intracellular acidification and cAMP levels. J Gen Microbiol 136: 2537-2543. doi: 10.1099/00221287-136-12-2537
|
| [254] | Bouillet LE, Cardoso AS, Perovano E, et al. (2011) The involvement of calcium carriers and of the vacuole in the glucose-induced calcium signaling and activation of the plasma membrane H+-ATPase in Saccharomyces cerevisiae cells. Cell Calcium 51: 72-81. |
| [255] |
Tr��pia M, Cardoso A, Tisi R, et al. (2006) Calcium signaling and sugar-induced activation of plasma membrane H+-ATPase in Saccharomyces cerevisiae cells. Biochem Biophys Res Commun 343: 1234-1243. doi: 10.1016/j.bbrc.2006.03.078
|
| [256] |
Zhang Y, Rao R (2008) A spoke in the wheel: calcium spikes disrupt yeast cell cycle. Cell Cycle 7: 870-873. doi: 10.4161/cc.7.7.5616
|
| [257] |
Burgoyne R, Clague M (2003) Calcium and calmodulin in membrane fusion. Biochim Biophys Acta 1641: 137-143. doi: 10.1016/S0167-4889(03)00089-2
|
| [258] |
Garrett-Engele P, Moilanen B, Cyert M (1995) Calcineurin, the Ca2+/calmodulin-dependent protein phosphatase, is essential in yeast mutants with cell integrity defects and in mutants that lack a functional vacuolar H+-ATPase. Mol Cell Biol 15: 4103-4114. doi: 10.1128/MCB.15.8.4103
|
| [259] |
Deka R, Kumar R, Tamuli R (2011) Neurospora crassa homologue of Neuronal Calcium Sensor-1 has a role in growth, calcium stress tolerance, and ultraviolet survival. Genetica 139: 885-894. doi: 10.1007/s10709-011-9592-y
|
| [260] |
Deka R, Tamuli R (2013) Neurospora crassa ncs-1, mid-1 and nca-2 double-mutant phenotypes suggest diverse interaction among three Ca2+-regulating gene products. J Genet 92: 559-563. doi: 10.1007/s12041-013-0270-y
|
| [261] |
Tamuli R, Kumar R, Deka R (2011) Cellular roles of neuronal calcium sensor-1 and calcium/calmodulin-dependent kinases in fungi. J Basic Microbiol 51: 120-128. doi: 10.1002/jobm.201000184
|
| [262] |
Barman A, Tamuli R (2015) Multiple cellular roles of Neurospora crassa plc-1, splA2, and cpe-1 in regulation of cytosolic free calcium, carotenoid accumulation, stress responses, and acquisition of thermotolerance. J Microbiol 53: 226-235. doi: 10.1007/s12275-015-4465-1
|
| [263] |
Tamuli R, Deka R, Borkovich KA (2016) Calcineurin Subunits A and B Interact to Regulate Growth and Asexual and Sexual Development in Neurospora crassa. PLoS One 11: e0151867. doi: 10.1371/journal.pone.0151867
|
| [264] | Davenport K, Sohaskey M, Kamada Y, et al. (1995) A second osmosensing signal transduction pathway in yeast. Hypotonic shock activates the PKC1 protein kinase-regulated cell integrity pathway. J Biol Chem 270: 30157-30161. |
| [265] |
Mizunuma M, Hirata D, Miyahara K, et al. (1998) Role of calcineurin and Mpk1 in regulating the onset of mitosis in budding yeast. Nature 392: 303-306. doi: 10.1038/32695
|
| [266] |
Levin DE (2005) Cell wall integrity signaling in Saccharomyces cerevisiae. Microbiol Mol Biol Rev 69: 262-291. doi: 10.1128/MMBR.69.2.262-291.2005
|
| [267] | Nakamura T, Ohmoto T, Hirata D, et al. (1996) Genetic evidence for the functional redundancy of the calcineurin- and Mpk1-mediated pathways in the regulation of cellular events important for growth in Saccharomyces cerevisiae. Mol Gen Genet 251: 211-219. |
| [268] |
Mazur P, Morin N, Baginsky W, et al. (1995) Differential expression and function of two homologous subunits of yeast 1,3-beta-D-glucan synthase. Mol Cell Biol 15: 5671-5681. doi: 10.1128/MCB.15.10.5671
|
| [269] |
Wang X, Sheff MA, Simpson DM, et al. (2011) Ste11p MEKK signals through HOG, mating, calcineurin and PKC pathways to regulate the FKS2 gene. BMC Mol Biol 12: 51. doi: 10.1186/1471-2199-12-51
|
| [270] |
Serrano R, Ruiz A, Bernal D, et al. (2002) The transcriptional response to alkaline pH in Saccharomyces cerevisiae: evidence for calcium-mediated signalling. Mol Microbiol 46: 1319-1333. doi: 10.1046/j.1365-2958.2002.03246.x
|
| [271] |
Lavina WA, Hermansyah, Sugiyama M, et al. (2013) Functionally redundant protein phosphatase genes PTP2 and MSG5 co-regulate the calcium signaling pathway in Saccharomyces cerevisiae upon exposure to high extracellular calcium concentration. J Biosci Bioeng 115: 138-146. doi: 10.1016/j.jbiosc.2012.08.022
|
| [272] |
Lagorce A, Hauser NC, Labourdette D, et al. (2003) Genome-wide analysis of the response to cell wall mutations in the yeast Saccharomyces cerevisiae. J Biol Chem 278: 20345-20357. doi: 10.1074/jbc.M211604200
|
| [273] |
Colabardini AC, Ries LN, Brown NA, et al. (2014) Protein kinase C overexpression suppresses calcineurin-associated defects in Aspergillus nidulans and is involved in mitochondrial function. PLoS One 9: e104792. doi: 10.1371/journal.pone.0104792
|
| [274] |
Teepe AG, Loprete DM, He Z, et al. (2007) The protein kinase C orthologue PkcA plays a role in cell wall integrity and polarized growth in Aspergillus nidulans. Fungal Genet Biol 44: 554-562. doi: 10.1016/j.fgb.2006.10.001
|
| [275] |
Herrmann M, Sprote P, Brakhage AA (2006) Protein kinase C (PkcA) of Aspergillus nidulans is involved in penicillin production. Appl Environ Microbiol 72: 2957-2970. doi: 10.1128/AEM.72.4.2957-2970.2006
|
| [276] |
Ichinomiya M, Uchida H, Koshi Y, et al. (2007) A protein kinase C-encoding gene, pkcA, is essential to the viability of the filamentous fungus Aspergillus nidulans. Biosci Biotechnol Biochem 71: 2787-2799. doi: 10.1271/bbb.70409
|
| [277] |
Ronen R, Sharon H, Levdansky E, et al. (2007) The Aspergillus nidulans pkcA gene is involved in polarized growth, morphogenesis and maintenance of cell wall integrity. Curr Genet 51: 321-329. doi: 10.1007/s00294-007-0129-y
|
| [278] |
Colabardini AC, De Castro PA, De Gouvea PF, et al. (2010) Involvement of the Aspergillus nidulans protein kinase C with farnesol tolerance is related to the unfolded protein response. Mol Microbiol 78: 1259-1279. doi: 10.1111/j.1365-2958.2010.07403.x
|
| [279] |
Katayama T, Uchida H, Ohta A, et al. (2012) Involvement of protein kinase C in the suppression of apoptosis and in polarity establishment in Aspergillus nidulans under conditions of heat stress. PLoS One 7: e50503. doi: 10.1371/journal.pone.0050503
|
| [280] |
Schumacher J, Viaud M, Simon A, et al. (2008) The Galpha subunit BCG1, the phospholipase C (BcPLC1) and the calcineurin phosphatase co-ordinately regulate gene expression in the grey mould fungus Botrytis cinerea. Mol Microbiol 67: 1027-1050. doi: 10.1111/j.1365-2958.2008.06105.x
|
| [281] |
Lev S, Desmarini D, Li C, et al. (2013) Phospholipase C of Cryptococcus neoformans regulates homeostasis and virulence by providing inositol trisphosphate as a substrate for Arg1 kinase. Infect Immun 81: 1245-1255. doi: 10.1128/IAI.01421-12
|
| [282] |
Galagan JE, Calvo SE, Borkovich KA, et al. (2003) The genome sequence of the filamentous fungus Neurospora crassa. Nature 422: 859-868. doi: 10.1038/nature01554
|
| [283] |
Borkovich KA, Alex LA, Yarden O, et al. (2004) Lessons from the genome sequence of Neurospora crassa: tracing the path from genomic blueprint to multicellular organism. Microbiol Mol Biol Rev 68: 1-108. doi: 10.1128/MMBR.68.1.1-108.2004
|
| [284] |
Silverman-Gavrila LB, Lew RR (2002) An IP3-activated Ca2+ channel regulates fungal tip growth. J Cell Sci 115: 5013-5025. doi: 10.1242/jcs.00180
|
| [285] |
Silverman-Gavrila LB, Lew RR (2003) Calcium gradient dependence of Neurospora crassa hyphal growth. Microbiology 149: 2475-2485. doi: 10.1099/mic.0.26302-0
|
| [286] |
Silverman-Gavrila LB, Lew RR (2001) Regulation of the tip-high [Ca2+] gradient in growing hyphae of the fungus Neurospora crassa. Eur J Cell Biol 80: 379-390. doi: 10.1078/0171-9335-00175
|
| [287] |
Rho HS, Jeon J, Lee YH (2009) Phospholipase C-mediated calcium signalling is required for fungal development and pathogenicity in Magnaporthe oryzae. Mol Plant Pathol 10: 337-346. doi: 10.1111/j.1364-3703.2009.00536.x
|
| [288] |
Choi J, Kim KS, Rho HS, et al. (2011) Differential roles of the phospholipase C genes in fungal development and pathogenicity of Magnaporthe oryzae. Fungal Genet Biol 48: 445-455. doi: 10.1016/j.fgb.2011.01.001
|
| [289] |
Flick JS, Thorner J (1993) Genetic and biochemical characterization of a phosphatidylinositol-specific phospholipase C in Saccharomyces cerevisiae. Mol Cell Biol 13: 5861-5876. doi: 10.1128/MCB.13.9.5861
|
| [290] |
Payne W, Fitzgerald-Hayes M (1993) A mutation in PLC1, a candidate phosphoinositide-specific phospholipase C gene from Saccharomyces cerevisiae, causes aberrant mitotic chromosome segregation. Mol Cell Biol 13: 4351-4364. doi: 10.1128/MCB.13.7.4351
|
| [291] | DeLillo N, Romero C, Lin H, et al. (2003) Genetic evidence for a role of phospholipase C at the budding yeast kinetochore. Mol Genet Genomics 269: 261-270. |
| [292] |
Jun Y, Fratti R, Wickner W (2004) Diacylglycerol and its formation by phospholipase C regulate Rab- and SNARE-dependent yeast vacuole fusion. J Biol Chem 279: 53186-53195. doi: 10.1074/jbc.M411363200
|
| [293] |
Banfic H, Bedalov A, York JD, et al. (2013) Inositol pyrophosphates modulate S phase progression after pheromone-induced arrest in Saccharomyces cerevisiae. J Biol Chem 288: 1717-1725. doi: 10.1074/jbc.M112.412288
|
| [294] |
Coccetti P, Tisi R, Martegani E, et al. (1998) The PLC1 encoded phospholipase C in the yeast Saccharomyces cerevisiae is essential for glucose-induced phosphatidylinositol turnover and activation of plasma membrane H+-ATPase. Biochim Biophys Acta 1405: 147-154. doi: 10.1016/S0167-4889(98)00099-8
|
| [295] |
Nakajima-Shimada J, Iida H, Tsuji F, et al. (1991) Monitoring of intracellular calcium in Saccharomyces cerevisiae with an apoaequorin cDNA expression system. Proc Natl Acad Sci U S A 88: 6878-6882. doi: 10.1073/pnas.88.15.6878
|
| [296] |
Tisi R, Belotti F, Wera S, et al. (2004) Evidence for inositol triphosphate as a second messenger for glucose-induced calcium signalling in budding yeast. Curr Genet 45: 83-89. doi: 10.1007/s00294-003-0465-5
|
| [297] |
Cavero S, Traba J, Del Arco A, et al. (2005) The calcium-dependent ATP-Mg/Pi mitochondrial carrier is a target of glucose-induced calcium signalling in Saccharomyces cerevisiae. Biochem J. 392: 537-544. doi: 10.1042/BJ20050806
|
| [298] |
Bergsma J, Kasri N, Donaton M, et al. (2001) PtdIns(4,5)P(2) and phospholipase C-independent Ins(1,4,5)P(3) signals induced by a nitrogen source in nitrogen-starved yeast cells. Biochem J 359: 517-523. doi: 10.1042/bj3590517
|
| [299] |
Guillas I, Vernay A, Vitagliano JJ, et al. (2013) Phosphatidylinositol 4,5-bisphosphate is required for invasive growth in Saccharomyces cerevisiae. J Cell Sci 126: 3602-3614. doi: 10.1242/jcs.122606
|
| [300] | Rebecchi MJ, Pentyala SN (2000) Structure, function, and control of phosphoinositide-specific phospholipase C. Physiol Rev 80: 1291-1335. |
| [301] | Guiney EL, Goldman AR, Elias JE, et al. (2014) Calcineurin regulates the yeast synaptojanin Inp53/Sjl3 during membrane stress. Mol Biol Cell 26: 769-785. |
| [302] |
Ron D, Walter P (2007) Signal integration in the endoplasmic reticulum unfolded protein response. Nat Rev Mol Cell Biol 8: 519-529. doi: 10.1038/nrm2199
|
| [303] |
Mori K (2009) Signalling pathways in the unfolded protein response: development from yeast to mammals. J Biochem 146: 743-750. doi: 10.1093/jb/mvp166
|
| [304] |
Chen Y, Feldman DE, Deng C, et al. (2005) Identification of mitogen-activated protein kinase signaling pathways that confer resistance to endoplasmic reticulum stress in Saccharomyces cerevisiae. Mol Cancer Res 3: 669-677. doi: 10.1158/1541-7786.MCR-05-0181
|
| [305] |
Nikawa J, Yamashita S (1992) IRE1 encodes a putative protein kinase containing a membrane-spanning domain and is required for inositol phototrophy in Saccharomyces cerevisiae. Mol Microbiol 6: 1441-1446. doi: 10.1111/j.1365-2958.1992.tb00864.x
|
| [306] |
Nikawa J, Akiyoshi M, Hirata S, et al. (1996) Saccharomyces cerevisiae IRE2/HAC1 is involved in IRE1-mediated KAR2 expression. Nucleic Acids Res 24: 4222-4226. doi: 10.1093/nar/24.21.4222
|
| [307] |
Kim H, Kim A, Cunningham KW (2012) Vacuolar H+-ATPase (V-ATPase) promotes vacuolar membrane permeabilization and nonapoptotic death in stressed yeast. J Biol Chem 287: 19029-19039. doi: 10.1074/jbc.M112.363390
|
| [308] |
Kajiwara K, Muneoka T, Watanabe Y, et al. (2012) Perturbation of sphingolipid metabolism induces endoplasmic reticulum stress-mediated mitochondrial apoptosis in budding yeast. Mol Microbiol 86: 1246-1261. doi: 10.1111/mmi.12056
|
| [309] |
van Meer G, Voelker DR, Feigenson GW (2008) Membrane lipids: where they are and how they behave. Nat Rev Mol Cell Biol 9: 112-124. doi: 10.1038/nrm2330
|
| [310] |
Hannun YA, Obeid LM (2008) Principles of bioactive lipid signalling: lessons from sphingolipids. Nat Rev Mol Cell Biol 9: 139-150. doi: 10.1038/nrm2329
|
| [311] |
Jayaraman T, Marks AR (2000) Calcineurin is downstream of the inositol 1,4,5-trisphosphate receptor in the apoptotic and cell growth pathways. J Biol Chem 275: 6417-6420. doi: 10.1074/jbc.275.9.6417
|
| [312] |
Perrone GG, Tan SX, Dawes IW (2008) Reactive oxygen species and yeast apoptosis. Biochim Biophys Acta 1783: 1354-1368. doi: 10.1016/j.bbamcr.2008.01.023
|
| [313] |
Schroder M (2008) Engineering eukaryotic protein factories. Biotechnol Lett 30: 187-196. doi: 10.1007/s10529-007-9524-1
|
| [314] |
Giorgi C, De Stefani D, Bononi A, et al. (2009) Structural and functional link between the mitochondrial network and the endoplasmic reticulum. Int J Biochem Cell Biol 41: 1817-1827. doi: 10.1016/j.biocel.2009.04.010
|
| [315] |
Cebulski J, Malouin J, Pinches N, et al. (2011) Yeast Bax inhibitor, Bxi1p, is an ER-localized protein that links the unfolded protein response and programmed cell death in Saccharomyces cerevisiae. PLoS One 6: e20882. doi: 10.1371/journal.pone.0020882
|
| [316] |
Alby K, Schaefer D, Sherwood RK, et al. (2010) Identification of a cell death pathway in Candida albicans during the response to pheromone. Eukaryot Cell 9: 1690-1701. doi: 10.1128/EC.00155-10
|
| [317] |
Lu H, Zhu Z, Dong L, et al. (2011) Lack of trehalose accelerates H2O2-induced Candida albicans apoptosis through regulating Ca2+ signaling pathway and caspase activity. PLoS One 6: e15808. doi: 10.1371/journal.pone.0015808
|
| [318] |
Hwang JH, Choi H, Kim AR, et al. (2014) Hibicuslide C-induced cell death in Candida albicans involves apoptosis mechanism. J Appl Microbiol 117: 1400-1411. doi: 10.1111/jam.12633
|
| [319] |
Gupta S, Ton V, Beaudry V, et al. (2003) Antifungal activity of amiodarone is mediated by disruption of calcium homeostasis. J Biol Chem 278: 28831-28839. doi: 10.1074/jbc.M303300200
|
| [320] |
Pozniakovsky AI, Knorre DA, Markova OV, et al. (2005) Role of mitochondria in the pheromone- and amiodarone-induced programmed death of yeast. J Cell Biol 168: 257-269. doi: 10.1083/jcb.200408145
|
| [321] | Zheng K, Pan J, Ye L, et al. (2007) Programmed cell death-involved aluminum toxicity in yeast alleviated by antiapoptotic members with decreased calcium signals. Plant Physiol 143: 38-49. |
| [322] |
Pereira C, Silva RD, Saraiva L, et al. (2008) Mitochondria-dependent apoptosis in yeast. Biochim Biophys Acta 1783: 1286-1302. doi: 10.1016/j.bbamcr.2008.03.010
|
| [323] | Lauff DB, Santa-Maria GE (2010) Potassium deprivation is sufficient to induce a cell death program in Saccharomyces cerevisiae. FEMS Yeast Res 10: 497-507. |
| [324] |
Gunter TE, Sheu SS (2009) Characteristics and possible functions of mitochondrial Ca2+ transport mechanisms. Biochim Biophys Acta 1787: 1291-1308. doi: 10.1016/j.bbabio.2008.12.011
|
| [325] |
Chinopoulos C, Adam-Vizi V (2010) Mitochondrial Ca2+ sequestration and precipitation revisited. FEBS J 277: 3637-3651. doi: 10.1111/j.1742-4658.2010.07755.x
|
| [326] |
Williams GS, Boyman L, Chikando AC, et al. (2013) Mitochondrial calcium uptake. Proc Natl Acad Sci U S A 110: 10479-10486. doi: 10.1073/pnas.1300410110
|
| [327] |
Virag A, Harris SD (2006) The Spitzenkorper: a molecular perspective. Mycol Res 110: 4-13. doi: 10.1016/j.mycres.2005.09.005
|
| [328] |
Muend S, Rao R (2008) Fungicidal activity of amiodarone is tightly coupled to calcium influx. FEMS Yeast Res 8: 425-431. doi: 10.1111/j.1567-1364.2008.00354.x
|
| [329] |
Roberts SK, McAinsh M, Widdicks L (2012) Cch1p mediates Ca2+ influx to protect Saccharomyces cerevisiae against eugenol toxicity. PLoS One 7: e43989. doi: 10.1371/journal.pone.0043989
|
| [330] |
Imura A, Tsuji Y, Murata M, et al. (2007) ��-Klotho as a Regulator of Calcium Homeostasis. Science 316: 1615-1618. doi: 10.1126/science.1135901
|
| [331] | Yokoyama H, Mizunuma M, Okamoto M, et al. (2006) Involvement of calcineurin-dependent degradation of Yap1p in Ca2+-induced G2 cell-cycle regulation in Saccharomyces cerevisiae. EMBO Rep 7: 519-524. |
| [332] |
Tsubakiyama R, Mizunuma M, Gengyo A, et al. (2011) Implication of Ca2+ in the regulation of replicative life span of budding yeast. J Biol Chem 286: 28681-28687. doi: 10.1074/jbc.M111.231415
|
| [333] |
Maeta K, Izawa S, Inoue Y (2005) Methylglyoxal, a metabolite derived from glycolysis, functions as a signal initiator of the high osmolarity glycerol-mitogen-activated protein kinase cascade and calcineurin/Crz1-mediated pathway in Saccharomyces cerevisiae. J Biol Chem 280: 253-260. doi: 10.1074/jbc.M408061200
|
| [334] | Shitamukai A, Hirata D, Sonobe S, et al. (2004) Evidence for antagonistic regulation of cell growth by the calcineurin and high osmolarity glycerol pathways in Saccharomyces cerevisiae. J Biol Chem 279: 3651-3661. |
| [335] |
Kullas AL, Martin SJ, Davis D (2007) Adaptation to environmental pH: integrating the Rim101 and calcineurin signal transduction pathways. Mol Microbiol 66: 858-871. doi: 10.1111/j.1365-2958.2007.05929.x
|
| [336] |
Lengeler KB, Davidson RC, D'Souza C, et al. (2000) Signal transduction cascades regulating fungal development and virulence. Microbiol Mol Biol Rev 64: 746-785. doi: 10.1128/MMBR.64.4.746-785.2000
|
| [337] |
Marchetti O, Entenza JM, Sanglard D, et al. (2000) Fluconazole plus cyclosporine: a fungicidal combination effective against experimental endocarditis due to Candida albicans. Antimicrob Agents Chemother 44: 2932-2938. doi: 10.1128/AAC.44.11.2932-2938.2000
|
| [338] |
Marchetti O, Moreillon P, Glauser MP, et al. (2000) Potent synergism of the combination of fluconazole and cyclosporine in Candida albicans. Antimicrob Agents Chemother 44: 2373-2381. doi: 10.1128/AAC.44.9.2373-2381.2000
|
| [339] |
Reedy JL, Filler SG, Heitman J (2010) Elucidating the Candida albicans calcineurin signaling cascade controlling stress response and virulence. Fungal Genet Biol 47: 107-116. doi: 10.1016/j.fgb.2009.09.002
|
| [340] |
Miyazaki T, Yamauchi S, Inamine T, et al. (2010) Roles of calcineurin and Crz1 in antifungal susceptibility and virulence of Candida glabrata. Antimicrob Agents Chemother 54: 1639-1643. doi: 10.1128/AAC.01364-09
|
| [341] |
Seider K, Gerwien F, Kasper L, et al. (2014) Immune evasion, stress resistance, and efficient nutrient acquisition are crucial for intracellular survival of Candida glabrata within macrophages. Eukaryot Cell 13: 170-183. doi: 10.1128/EC.00262-13
|
| [342] |
Odom A, Muir S, Lim E, et al. (1997) Calcineurin is required for virulence of Cryptococcus neoformans. EMBO J 16: 2576-2589. doi: 10.1093/emboj/16.10.2576
|
| [343] |
Liu S, Hou Y, Liu W, et al. (2015) Components of the calcium-calcineurin signaling pathway in fungal cells and their potential as antifungal targets. Eukaryot Cell 14: 324-334. doi: 10.1128/EC.00271-14
|
| [344] | Blankenship JR, Steinbach WJ, Perfect JR, et al. (2003) Teaching old drugs new tricks: reincarnating immunosuppressants as antifungal drugs. Curr Opin Investig Drugs 4: 192-199. |
| [345] |
Cruz MC, Del Poeta M, Wang P, et al. (2000) Immunosuppressive and nonimmunosuppressive cyclosporine analogs are toxic to the opportunistic fungal pathogen Cryptococcus neoformans via cyclophilin-dependent inhibition of calcineurin. Antimicrob Agents Chemother 44: 143-149. doi: 10.1128/AAC.44.1.143-149.2000
|
| [346] |
Cruz MC, Goldstein AL, Blankenship J, et al. (2001) Rapamycin and less immunosuppressive analogs are toxic to Candida albicans and Cryptococcus neoformans via FKBP12-dependent inhibition of TOR. Antimicrob Agents Chemother 45: 3162-3170. doi: 10.1128/AAC.45.11.3162-3170.2001
|
| [347] |
Rodr��guez A, Roy J, Mart��nez-Mart��nez S, et al. (2009) A conserved docking surface on calcineurin mediates interaction with substrates and immunosuppressants. Mol Cell 33: 616-626. doi: 10.1016/j.molcel.2009.01.030
|
| [348] |
Edlind T, Smith L, Henry K, et al. (2002) Antifungal activity in Saccharomyces cerevisiae is modulated by calcium signalling. Mol Microbiol 46: 257-268. doi: 10.1046/j.1365-2958.2002.03165.x
|
| [349] |
Maesaki S, Marichal P, Hossain MA, et al. (1998) Synergic effects of tactolimus and azole antifungal agents against azole-resistant Candida albicans strains. J Antimicrob Chemother 42: 747-753. doi: 10.1093/jac/42.6.747
|
| [350] |
Steinbach WJ, Schell WA, Blankenship JR, et al. (2004) In vitro interactions between antifungals and immunosuppressants against Aspergillus fumigatus. Antimicrob Agents Chemother 48: 1664-1669. doi: 10.1128/AAC.48.5.1664-1669.2004
|
| [351] |
Schumacher J, de Larrinoa IF, Tudzynski B (2008) Calcineurin-responsive zinc finger transcription factor CRZ1 of Botrytis cinerea is required for growth, development, and full virulence on bean plants. Eukaryot Cell 7: 584-601. doi: 10.1128/EC.00426-07
|
| [352] |
Onyewu C, Wormley FL Jr, Perfect JR, et al. (2004) The calcineurin target, Crz1, functions in azole tolerance but is not required for virulence of Candida albicans. Infect Immun 72: 7330-7333. doi: 10.1128/IAI.72.12.7330-7333.2004
|
| [353] | Nakamura T, Takahashi H (2006) Epidemiological study of Candida infections in blood: susceptibilities of Candida spp. to antifungal agents, and clinical features associated with the candidemia. J Infect Chemother 12: 132-138. |
| [354] | Rogers TR (2002) Antifungal drug resistance: does it matter? Int J Infect Dis 6: S47-53. |
| [355] | Berman J, Sudbery PE (2002) Candida albicans: a molecular revolution built on lessons from budding yeast. Nat Rev Genet 3: 918-930. |
| [356] |
Lopez-Ribot JL (2005) Candida albicans biofilms: more than filamentation. Curr Biol 15: R453-455. doi: 10.1016/j.cub.2005.06.020
|
| [357] | Lo HJ, K?hler JR, DiDomenico B, et al. (1997) Nonfilamentous C. albicans mutants are avirulent. Cell 90: 939-949. |
| [358] |
Douglas LJ (2003) Candida biofilms and their role in infection. Trends Microbiol 11: 30-36. doi: 10.1016/S0966-842X(02)00002-1
|
| [359] | Hawser SP, Douglas LJ (1994) Biofilm formation by Candida species on the surface of catheter materials in vitro. Infect Immun 62: 915-921. |
| [360] |
Liu H (2001) Transcriptional control of dimorphism in Candida albicans. Curr Opin Microbiol 4: 728-735. doi: 10.1016/S1369-5274(01)00275-2
|
| [361] |
Nobile CJ, Fox EP, Nett JE, et al. (2012) A recently evolved transcriptional network controls biofilm development in Candida albicans. Cell 148: 126-138. doi: 10.1016/j.cell.2011.10.048
|
| [362] |
Brown AJ, Gow NA (1999) Regulatory networks controlling Candida albicans morphogenesis. Trends Microbiol 7: 333-338. doi: 10.1016/S0966-842X(99)01556-5
|
| [363] |
Sabie FT, Gadd GM (1989) Involvement of a Ca2+-calmodulin interaction in the yeast-mycelial (Y-M) transition of Candida albicans. Mycopathologia 108: 47-54. doi: 10.1007/BF00436783
|
| [364] |
Blankenship JR, Wormley FL, Boyce MK, et al. (2003) Calcineurin is essential for Candida albicans survival in serum and virulence. Eukaryot Cell 2: 422-430. doi: 10.1128/EC.2.3.422-430.2003
|
| [365] |
Blankenship JR, Heitman J (2005) Calcineurin is required for Candida albicans to survive calcium stress in serum. Infect Immun 73: 5767-5774. doi: 10.1128/IAI.73.9.5767-5774.2005
|
| [366] |
Sanglard D, Ischer F, Marchetti O, et al. (2003) Calcineurin A of Candida albicans: involvement in antifungal tolerance, cell morphogenesis and virulence. Mol Microbiol 48: 959-976. doi: 10.1046/j.1365-2958.2003.03495.x
|
| [367] |
Onyewu C, Blankenship JR, Del Poeta M, et al. (2003) Ergosterol biosynthesis inhibitors become fungicidal when combined with calcineurin inhibitors against Candida albicans, Candida glabrata, and Candida krusei. Antimicrob Agents Chemother 47: 956-964. doi: 10.1128/AAC.47.3.956-964.2003
|
| [368] |
Riscili BP, Wood KL (2009) Noninvasive pulmonary Aspergillus infections. Clin Chest Med 30: 315-335, vii. doi: 10.1016/j.ccm.2009.02.008
|
| [369] |
Lelievre L, Groh M, Angebault C, et al. (2013) Azole resistant Aspergillus fumigatus: an emerging problem. Med Mal Infect 43: 139-145. doi: 10.1016/j.medmal.2013.02.010
|
| [370] |
Denning DW, Perlin DS (2011) Azole resistance in Aspergillus: a growing public health menace. Future Microbiol 6: 1229-1232. doi: 10.2217/fmb.11.118
|
| [371] |
Todd RB, Davis MA, Hynes MJ (2007) Genetic manipulation of Aspergillus nidulans: heterokaryons and diploids for dominance, complementation and haploidization analyses. Nat Protoc 2: 822-830. doi: 10.1038/nprot.2007.113
|
| [372] |
Todd RB, Davis MA, Hynes MJ (2007) Genetic manipulation of Aspergillus nidulans: meiotic progeny for genetic analysis and strain construction. Nat Protoc 2: 811-821. doi: 10.1038/nprot.2007.112
|
| [373] |
Srikanta D, Santiago-Tirado FH, Doering TL (2014) Cryptococcus neoformans: historical curiosity to modern pathogen. Yeast 31: 47-60. doi: 10.1002/yea.2997
|
| [374] |
Perfect JR, Casadevall A (2002) Cryptococcosis. Infect Dis Clin North Am 16: 837-874. doi: 10.1016/S0891-5520(02)00036-3
|
| [375] |
Steenbergen JN, Casadevall A (2003) The origin and maintenance of virulence for the human pathogenic fungus Cryptococcus neoformans. Microbes Infect 5: 667-675. doi: 10.1016/S1286-4579(03)00092-3
|
| [376] |
Brown SM, Campbell LT, Lodge JK (2007) Cryptococcus neoformans, a fungus under stress. Curr Opin Microbiol 10: 320-325. doi: 10.1016/j.mib.2007.05.014
|
| [377] |
Bahn YS, Jung KW (2013) Stress signaling pathways for the pathogenicity of Cryptococcus. Eukaryot Cell 12: 1564-1577. doi: 10.1128/EC.00218-13
|
| [378] |
Fox DS, Cruz MC, Sia RA, et al. (2001) Calcineurin regulatory subunit is essential for virulence and mediates interactions with FKBP12-FK506 in Cryptococcus neoformans. Mol Microbiol 39: 835-849. doi: 10.1046/j.1365-2958.2001.02295.x
|
| [379] |
Kozubowski L, Aboobakar EF, Cardenas ME, et al. (2011) Calcineurin colocalizes with P-bodies and stress granules during thermal stress in Cryptococcus neoformans. Eukaryot Cell 10: 1396-1402. doi: 10.1128/EC.05087-11
|
| [380] |
Cruz MC, Fox DS, Heitman J (2001) Calcineurin is required for hyphal elongation during mating and haploid fruiting in Cryptococcus neoformans. EMBO J 20: 1020-1032. doi: 10.1093/emboj/20.5.1020
|
| [381] |
Fox DS, Heitman J (2002) Good fungi gone bad: the corruption of calcineurin. Bioessays 24: 894-903. doi: 10.1002/bies.10157
|
| [382] |
Kraus PR, Fox DS, Cox GM, et al. (2003) The Cryptococcus neoformans MAP kinase Mpk1 regulates cell integrity in response to antifungal drugs and loss of calcineurin function. Mol Microbiol 48: 1377-1387. doi: 10.1046/j.1365-2958.2003.03508.x
|
| [383] | Thompson JR, Douglas CM, Li W, et al. (1999) A glucan synthase FKS1 homolog in Cryptococcus neoformans is single copy and encodes an essential function. J Bacteriol 181: 444-453. |
| [384] |
Juvvadi PR, Gehrke C, Fortwendel JR, et al. (2013) Phosphorylation of Calcineurin at a novel serine-proline rich region orchestrates hyphal growth and virulence in Aspergillus fumigatus. PLoS Pathog 9: e1003564. doi: 10.1371/journal.ppat.1003564
|
| [385] |
Bates S, MacCallum DM, Bertram G, et al. (2005) Candida albicans Pmr1p, a secretory pathway P-type Ca2+/Mn2+-ATPase, is required for glycosylation and virulence. J Biol Chem 280: 23408-23415. doi: 10.1074/jbc.M502162200
|
| [386] |
LaFayette SL, Collins C, Zaas AK, et al. (2010) PKC signaling regulates drug resistance of the fungal pathogen Candida albicans via circuitry comprised of Mkc1, calcineurin, and Hsp90. PLoS Pathog 6: e1001069. doi: 10.1371/journal.ppat.1001069
|
| [387] |
Yu Q, Ding X, Zhang B, et al. (2014) Inhibitory effect of verapamil on Candida albicans hyphal development, adhesion and gastrointestinal colonization. FEMS Yeast Res 14: 633-641. doi: 10.1111/1567-1364.12150
|
| [388] |
Ramage G, Wickes BL, Lopez-Ribot JL (2007) Inhibition on Candida albicans biofilm formation using divalent cation chelators (EDTA). Mycopathologia 164: 301-306. doi: 10.1007/s11046-007-9068-x
|
| [389] |
Raad II, Hachem RY, Hanna HA, et al. (2008) Role of ethylene diamine tetra-acetic acid (EDTA) in catheter lock solutions: EDTA enhances the antifungal activity of amphotericin B lipid complex against Candida embedded in biofilm. Int J Antimicrob Agents 32: 515-518. doi: 10.1016/j.ijantimicag.2008.06.020
|
| [390] |
Yu Q, Ding X, Xu N, et al. (2013) In vitro activity of verapamil alone and in combination with fluconazole or tunicamycin against Candida albicans biofilms. Int J Antimicrob Agents 41: 179-182. doi: 10.1016/j.ijantimicag.2012.10.009
|
| [391] |
Kmetzsch L, Staats CC, Cupertino JB, et al. (2013) The calcium transporter Pmc1 provides Ca2+ tolerance and influences the progression of murine cryptococcal infection. FEBS J 280: 4853-4864. doi: 10.1111/febs.12458
|
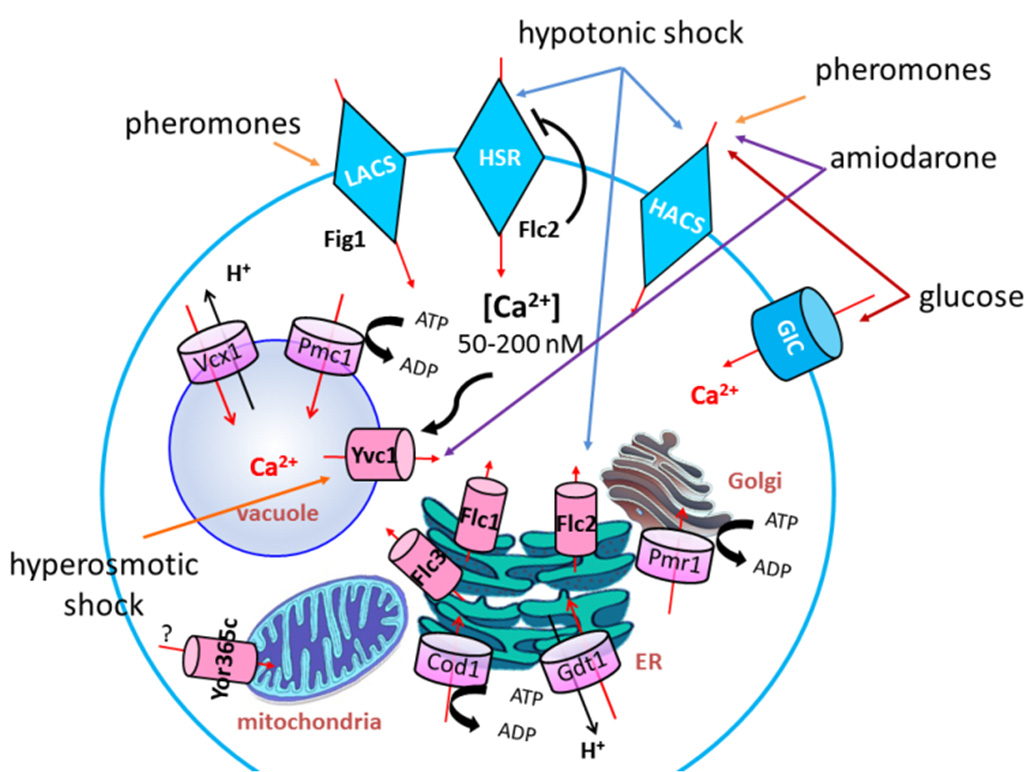
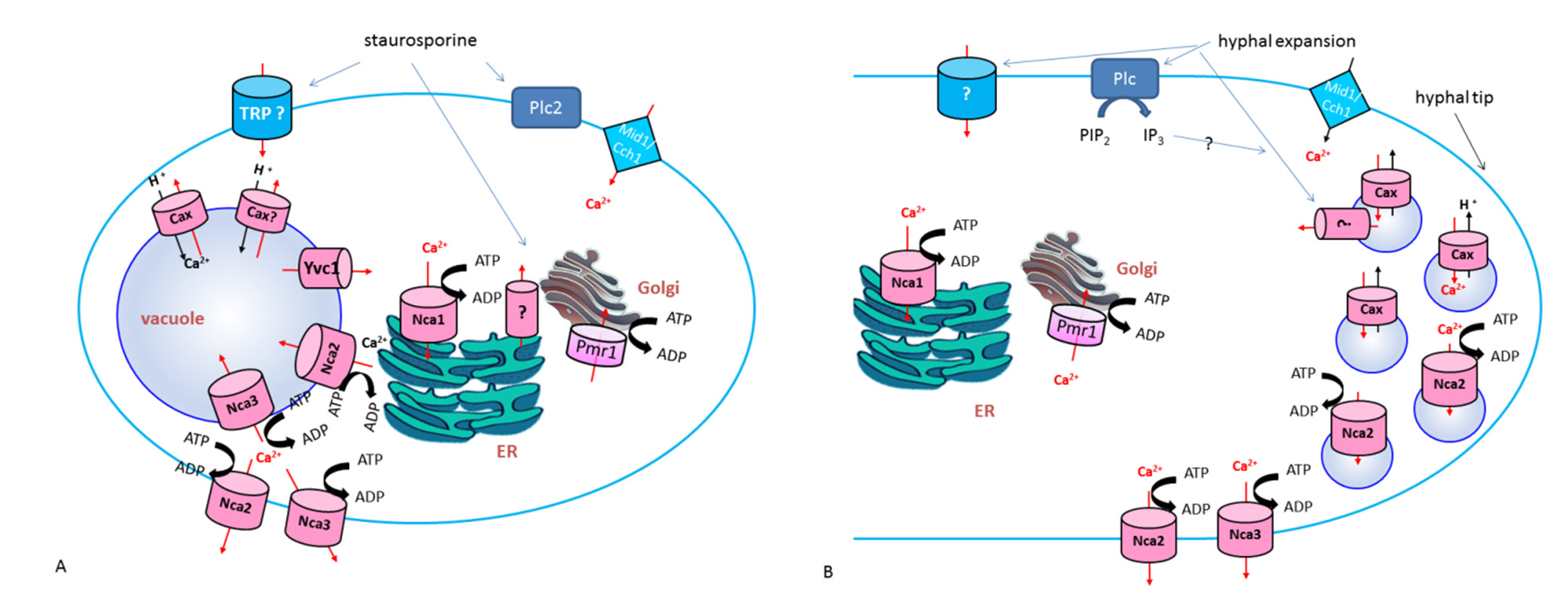
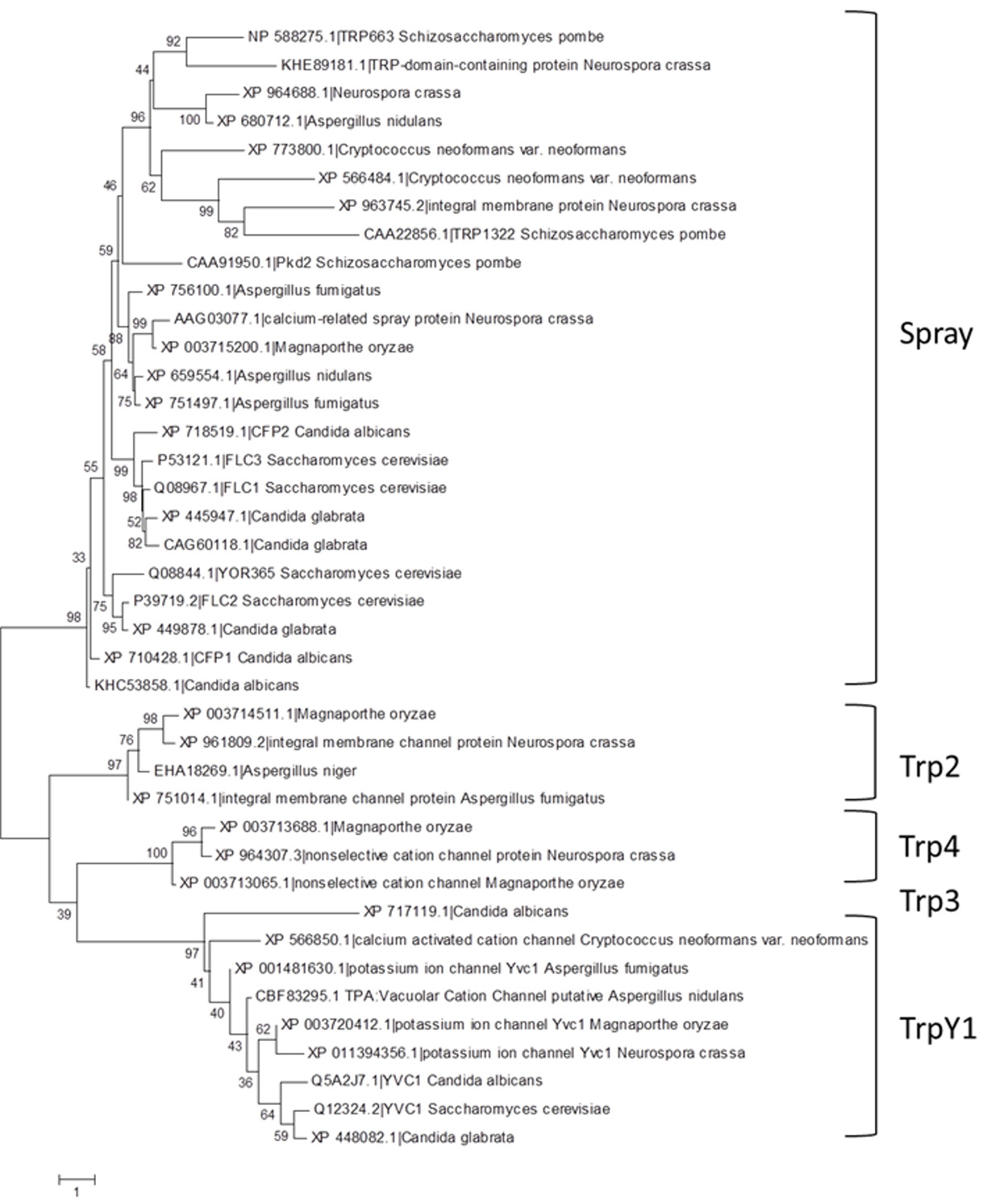
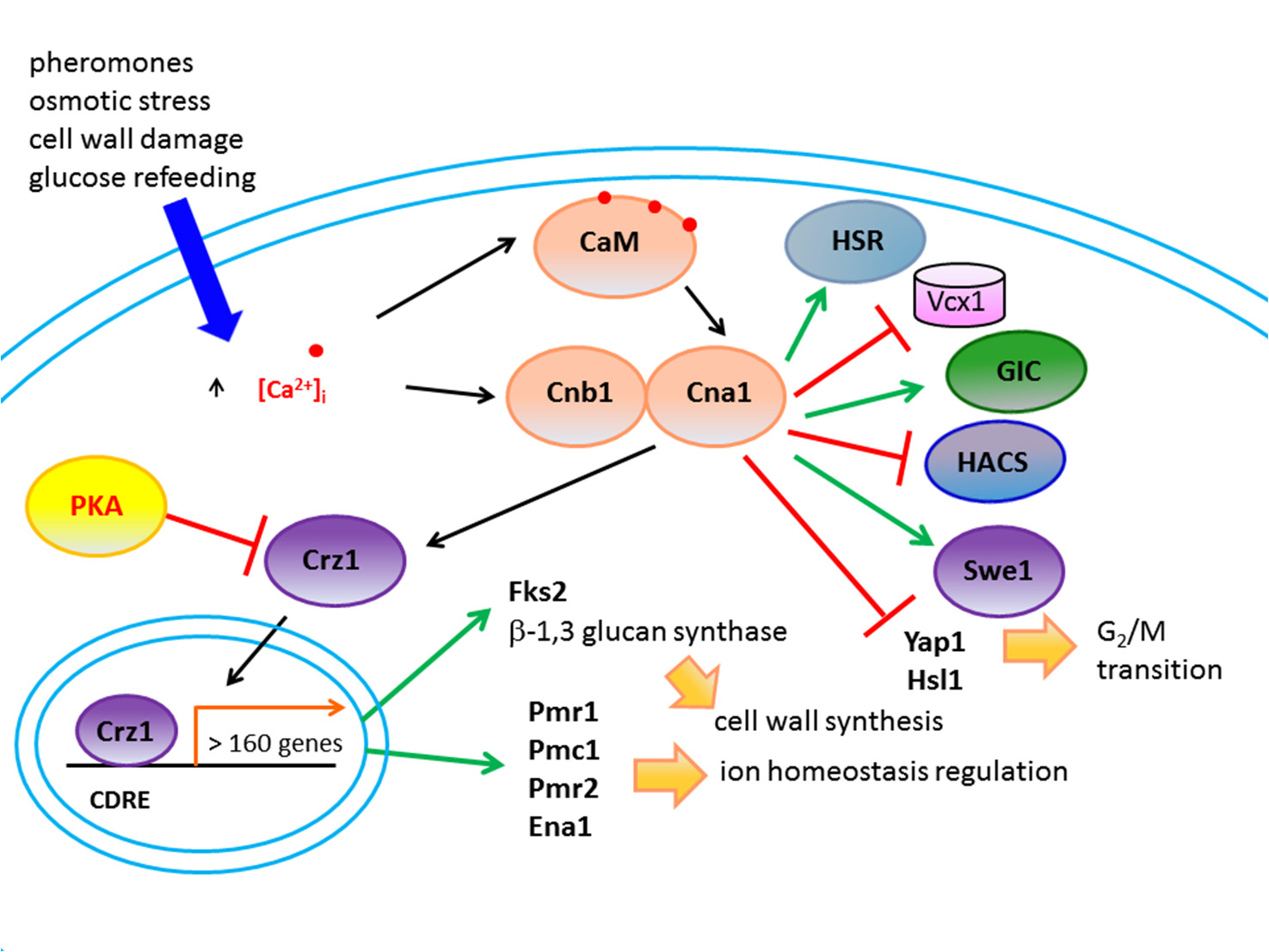
Figures(4) / Tables(1)

Renata Tisi, Marco Rigamonti, Silvia Groppi, Fiorella Belotti. Calcium homeostasis and signaling in fungi and their relevance for pathogenicity of yeasts and filamentous fungi[J]. AIMS Molecular Science, 2016, 3(4): 505-549. doi: 10.3934/molsci.2016.4.505







 DownLoad:
DownLoad: