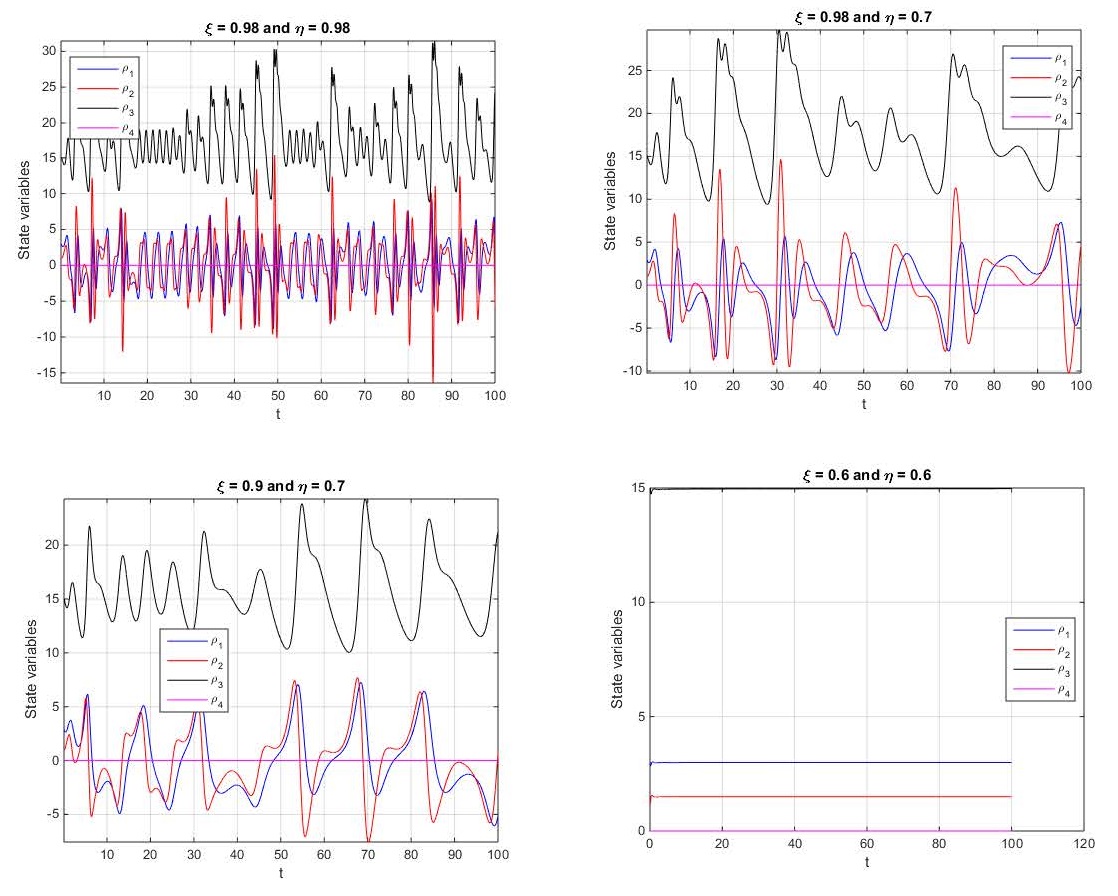
Although fractional calculus is about three centuries old, it has become the key to understanding many complex real-world phenomena. During the past few decades, many fractional derivatives have appeared. Among these, the fractal-fractional derivatives have shown acceptance in describing some real-world problems. In this paper, the Caputo, Atangana-Baleanu, and Caputo-Fabrizio fractal-fractional operators were applied to generate complex dynamics in a 4D dynamical system. Some conditions for the exact solutions' existence and uniqueness were demonstrated when the fractal-fractional operators are implemented into the mentioned 4D dynamical system. Some Ulam-Hyers stability results were demonstrated in the indicated fractal-fractional systems. Computation processes were carried out to demonstrate some graphical results that showed the existence of several complex dynamics in the considered system as the fractal-fractional operators are implemented. Furthermore, the computations of the system's Lyapunov exponents and the bifurcation diagrams were used to illustrate the wide range of chaotic dynamics that exist in the considered fractal-fractional 4D system. Existence of hidden chaotic attractors were also found. This interesting dynamical phenomenon was validated by the bifurcation diagrams and basin set of attraction.
Citation: A. E. Matouk. Chaos and hidden chaos in a 4D dynamical system using the fractal-fractional operators[J]. AIMS Mathematics, 2025, 10(3): 6233-6257. doi: 10.3934/math.2025284
Although fractional calculus is about three centuries old, it has become the key to understanding many complex real-world phenomena. During the past few decades, many fractional derivatives have appeared. Among these, the fractal-fractional derivatives have shown acceptance in describing some real-world problems. In this paper, the Caputo, Atangana-Baleanu, and Caputo-Fabrizio fractal-fractional operators were applied to generate complex dynamics in a 4D dynamical system. Some conditions for the exact solutions' existence and uniqueness were demonstrated when the fractal-fractional operators are implemented into the mentioned 4D dynamical system. Some Ulam-Hyers stability results were demonstrated in the indicated fractal-fractional systems. Computation processes were carried out to demonstrate some graphical results that showed the existence of several complex dynamics in the considered system as the fractal-fractional operators are implemented. Furthermore, the computations of the system's Lyapunov exponents and the bifurcation diagrams were used to illustrate the wide range of chaotic dynamics that exist in the considered fractal-fractional 4D system. Existence of hidden chaotic attractors were also found. This interesting dynamical phenomenon was validated by the bifurcation diagrams and basin set of attraction.

| [1] | G. Pertz, C. J. Gerhardt, Briefwechsel zwischen Leibniz, Hugens van Zulichem und dem Marquis de l'Hospital, Leibnizens gesammelte Werke, 1849. |
| [2] |
M. Caputo, Linear models of dissipation whose Q is almost frequency independent-II, Geophys. J. R. Astron. Soc., 13 (1967), 529–539. https://doi.org/10.1111/j.1365-246X.1967.tb02303.x doi: 10.1111/j.1365-246X.1967.tb02303.x

|
| [3] |
M. Caputo, M. Fabrizio, A new definition of fractional derivative without singular kernel, Prog. Fract. Differ. Appl., 1 (2015), 73. http://dx.doi.org/10.12785/pfda/010201 doi: 10.12785/pfda/010201
|
| [4] |
A. Atangana, D. Baleanu, Caputo-Fabrizio derivative applied to groundwater flow within confined aquifer, J. Eng. Mech., 143 (2017), D4016005. https://doi.org/10.1061/(ASCE)EM.1943-7889.0001091 doi: 10.1061/(ASCE)EM.1943-7889.0001091
|
| [5] |
A. Atangana, Fractal-fractional differentiation and integration: connecting fractal calculus and fractional calculus to predict complex system, Chaos Soliton. Fract., 102 (2017), 396–406. https://doi.org/10.1016/j.chaos.2017.04.027 doi: 10.1016/j.chaos.2017.04.027
|
| [6] | C. Sparrow, The Lorenz equations: Bifurcation, chaos and strange attractor, New York: Springer-Verlag, 1982. |
| [7] | A. Wolf, J. B. Swift, H. L. Swinney, J. A. Vastano, Determining Lyapunov exponents from a time series, Phys. D, 16 (1985) 285–317. |
| [8] | E. N. Lorenz, Deterministic non-periodic flow, J. Atmospheric Sci., 20 (1963), 130–141. |
| [9] | O. E. Rössler, An equation for continuous chaos, Phys. Lett. A, 57 (1976), 397–398. |
| [10] |
A. E. Matouk, H. N. Agiza, Bifurcations, chaos and synchronization in ADVP circuit with parallel resistor, J. Math. Anal. Appl., 341 (2008), 259–269. https://doi.org/10.1016/j.jmaa.2007.09.067 doi: 10.1016/j.jmaa.2007.09.067
|
| [11] |
A. E. Matouk, Dynamics and control in a novel hyperchaotic system, Int. J. Dyn. Cont., 7 (2019), 241. https://doi.org/10.1007/s40435-018-0439-6 doi: 10.1007/s40435-018-0439-6
|
| [12] |
I. Grigorenko, E. Grigorenko, Chaotic dynamics of the fractional Lorenz system, Phys. Rev. Lett., 91 (2003), 034101. https://doi.org/10.1103/PhysRevLett.91.034101 doi: 10.1103/PhysRevLett.91.034101
|
| [13] |
W. Zhang, S. Zhou, H. Li, H. Zhu, Chaos in a fractional-order Rössler system, Chaos Soliton. Fract., 42 (2009), 1684–1691. https://doi.org/10.1016/j.chaos.2009.03.069 doi: 10.1016/j.chaos.2009.03.069
|
| [14] |
A. E. Matouk, Chaos, feedback control and synchronization of a fractional-order modified autonomous Van der Pol-Duffing circuit, Commun. Nonlinear Sci. Numer. Simulat., 16 (2011), 975–986. https://doi.org/10.1016/j.cnsns.2010.04.027 doi: 10.1016/j.cnsns.2010.04.027
|
| [15] |
A. E. Matouk, A novel fractional-order system: Chaos, hyperchaos and applications to linear control, J. Appl. Comput. Mech., 7 (2021), 701–714. https://doi.10.22055/JACM.2020.35092.2561 doi: 10.22055/JACM.2020.35092.2561
|
| [16] |
S. Qureshi, A. Atangana, Fractal-fractional differentiation for the modeling and mathematical analysis of nonlinear diarrhea transmission dynamics under the use of real data, Chaos Soliton. Fract., 136 (2020), 109812. https://doi.org/10.1016/j.chaos.2020.109812 doi: 10.1016/j.chaos.2020.109812
|
| [17] |
A. Sami, A. Ali, R. Shafqat, N. Pakkaranang, M. Rahmamn, Analysis of food chain mathematical model under fractal fractional Caputo derivative, Math. Biosci. Eng., 20 (2022), 2094–2109. https://doi.org/10.3934/mbe.2023097 doi: 10.3934/mbe.2023097
|
| [18] |
N. Almutairi, S. Saber, H. Ahmad, The fractal-fractional Atangana-Baleanu operator for pneumonia disease: Stability, statistical and numerical analyses, AIMS Math., 8 (2023), 29382–29410. https://doi.org/10.3934/math.20231504 doi: 10.3934/math.20231504
|
| [19] |
M. Arif, P. Kumam, W. Kumam, A. Akgul, T. Sutthibutpong, Analysis of newly developed fractal-fractional derivative with power law kernel for MHD couple stress fluid in channel embedded in a porous medium, Sci. Rep., 11 (2021) 20858. https://doi.org/10.1038/s41598-021-00163-3 doi: 10.1038/s41598-021-00163-3
|
| [20] |
A. Khan, K. Shah, T. Abdeljawad, I. Amacha, Fractal fractional model for tuberculosis: existence and numerical solutions, Sci. Rep., 14 (2024) 12211. https://doi.org/10.1038/s41598-024-62386-4 doi: 10.1038/s41598-024-62386-4
|
| [21] |
K. Shah, T. Abdeljawad, On complex fractal-fractional order mathematical modeling of CO2 emanations from energy sector, Phys. Scripta, 99 (2024) 015226. https://doi.org/10.1088/1402-4896/ad1286 doi: 10.1088/1402-4896/ad1286
|
| [22] |
A. Dlamini, E. F. D. Goufo, M. Khumalo, On the Caputo-Fabrizio fractal fractional representation for the Lorenz chaotic system, AIMS Math., 6 (2021) 12395–12421. https://doi.org/10.3934/math.2021717 doi: 10.3934/math.2021717
|
| [23] |
I. Ul Haq, N. Ali, H. Ahmad, Analysis of a chaotic system using fractal-fractional derivatives with exponential decay type kernels, Math. Model. Control, 2 (2022) 185–199. https://doi.org/10.3934/mmc.2022019 doi: 10.3934/mmc.2022019
|
| [24] |
S. Saber, Control of chaos in the Burke-Shaw system of fractal-fractional order in the sense of Caputo-Fabrizio, J. Appl. Math. Comput. Mech., 23 (2024) 83–96. https://doi.org/10.17512/jamcm.2024.1.07 doi: 10.17512/jamcm.2024.1.07
|
| [25] | A. Granas, J. Dugundji, Fixed point theory, New York: Springer, 2005. |
| [26] |
A. Atangana, S. Qureshi, Modeling attractors of chaotic dynamical systems with fractal-fractional operators, Chaos Soliton. Fract., 123 (2019), 320–337. https://doi.org/10.1016/j.chaos.2019.04.020 doi: 10.1016/j.chaos.2019.04.020
|
| [27] |
N. V. Kuznetsov, G. Leonov, Hidden attractors in dynamical systems: systems with no equilibria, multistability and coexisting attractors, IFAC Proc., 47 (2014), 5445–5454. https://doi.org/10.3182/20140824-6-ZA-1003.02501 doi: 10.3182/20140824-6-ZA-1003.02501
|






















Figures(23)

A. E. Matouk. Chaos and hidden chaos in a 4D dynamical system using the fractal-fractional operators[J]. AIMS Mathematics, 2025, 10(3): 6233-6257. doi: 10.3934/math.2025284







 DownLoad:
DownLoad: