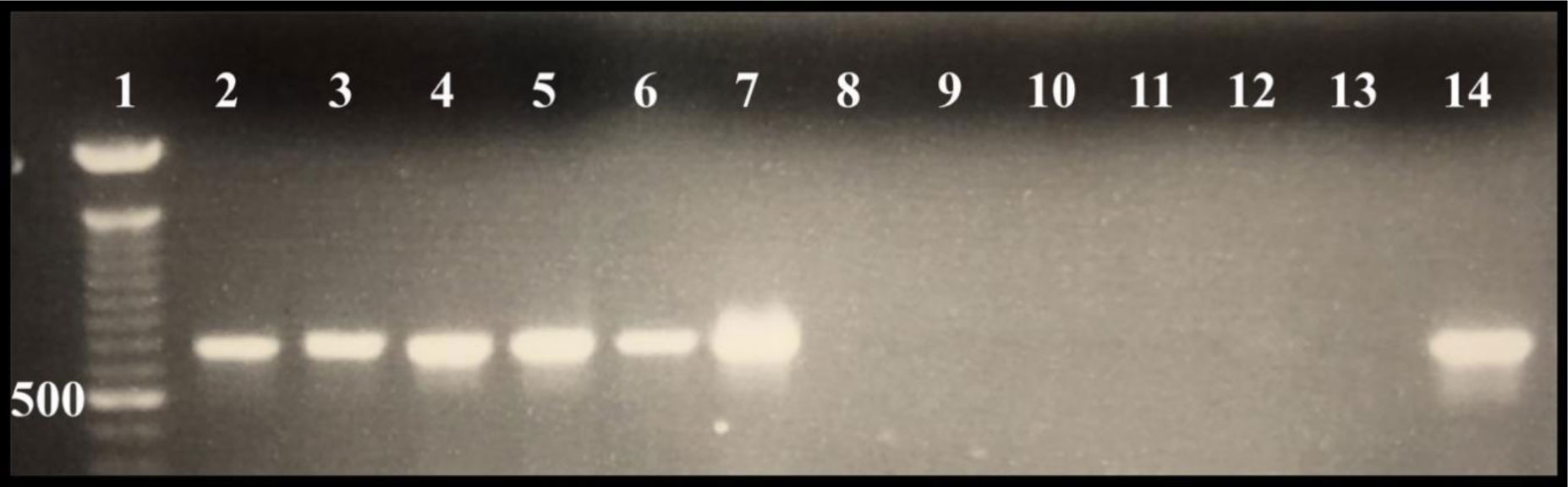
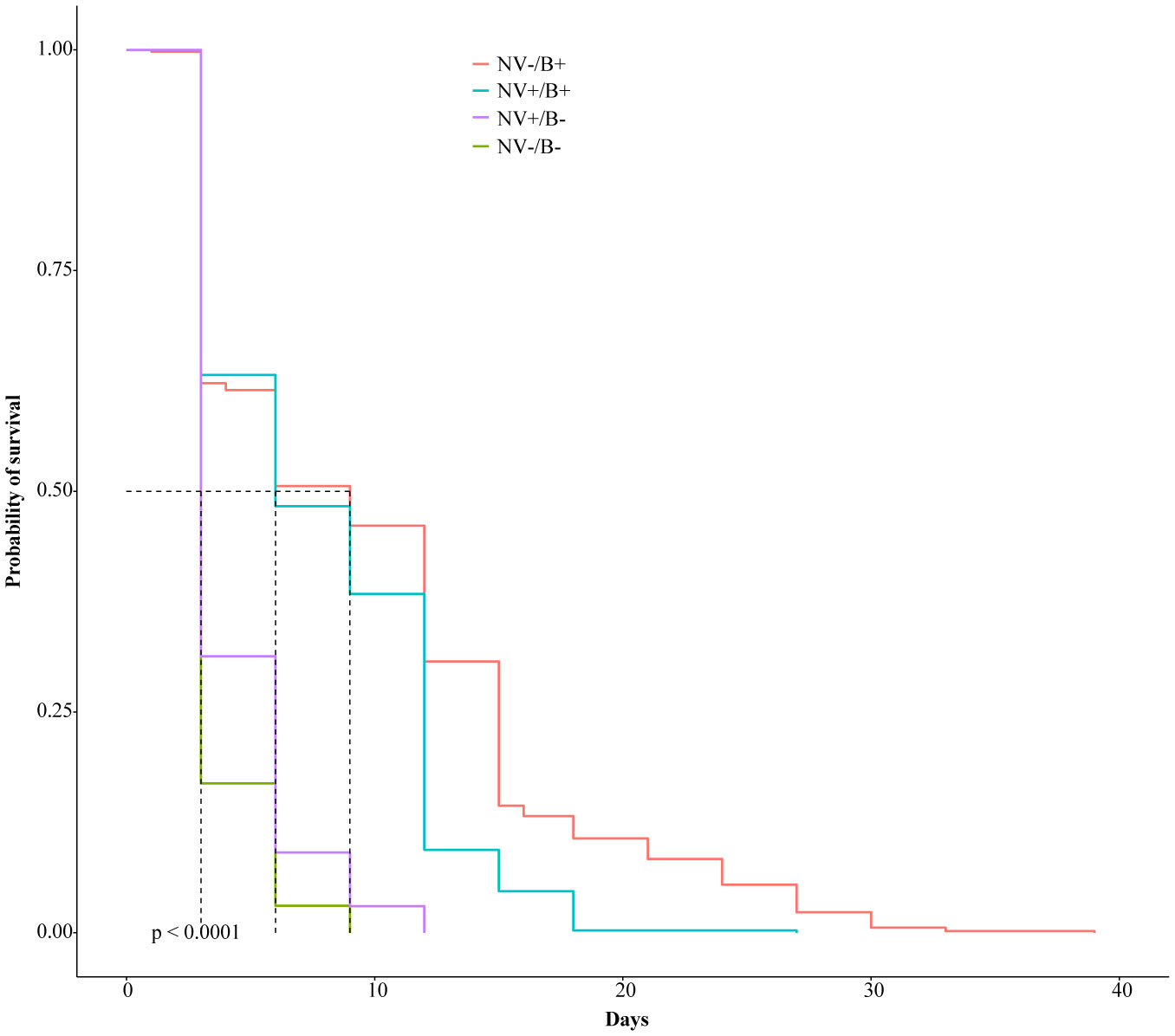
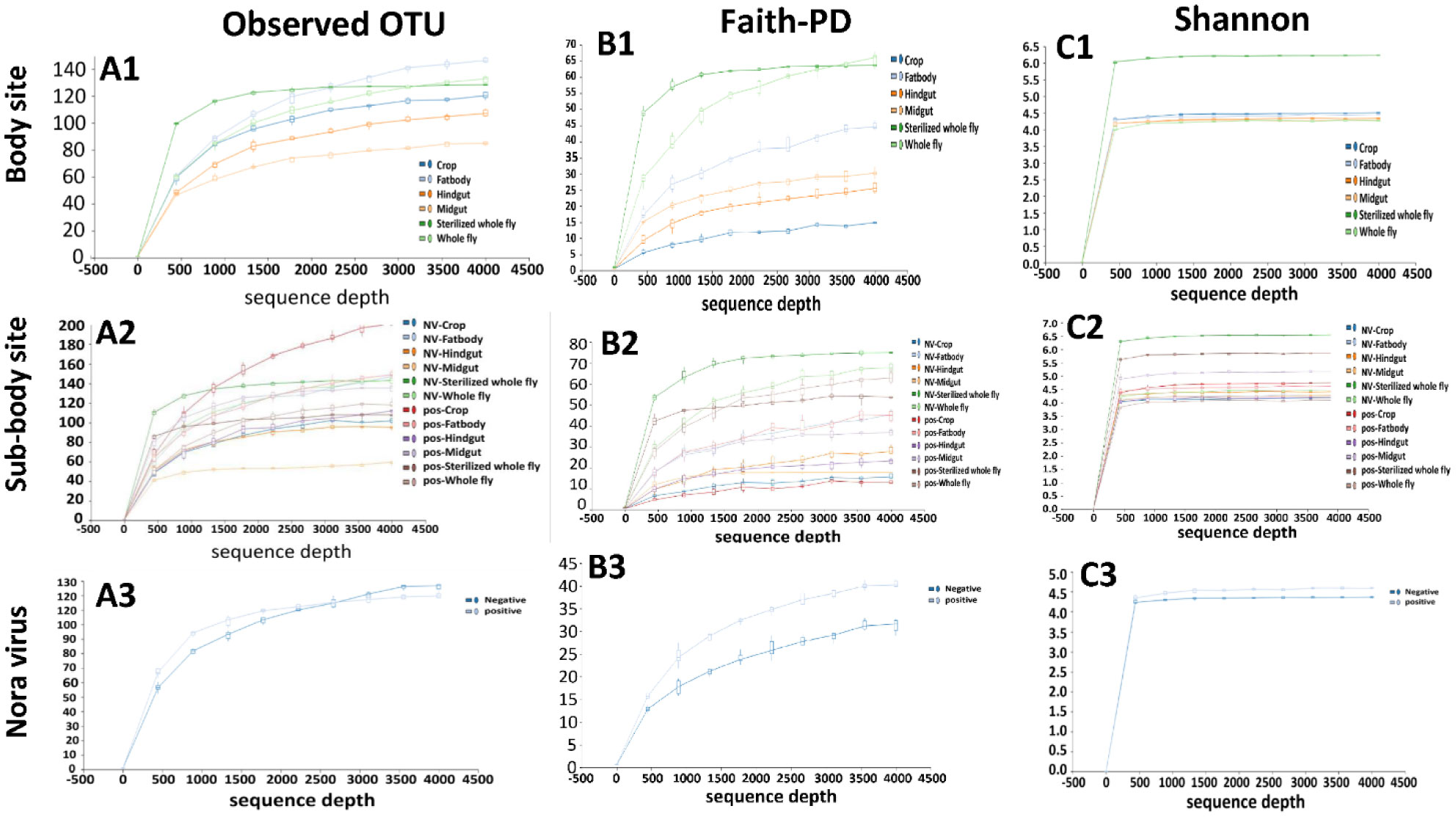
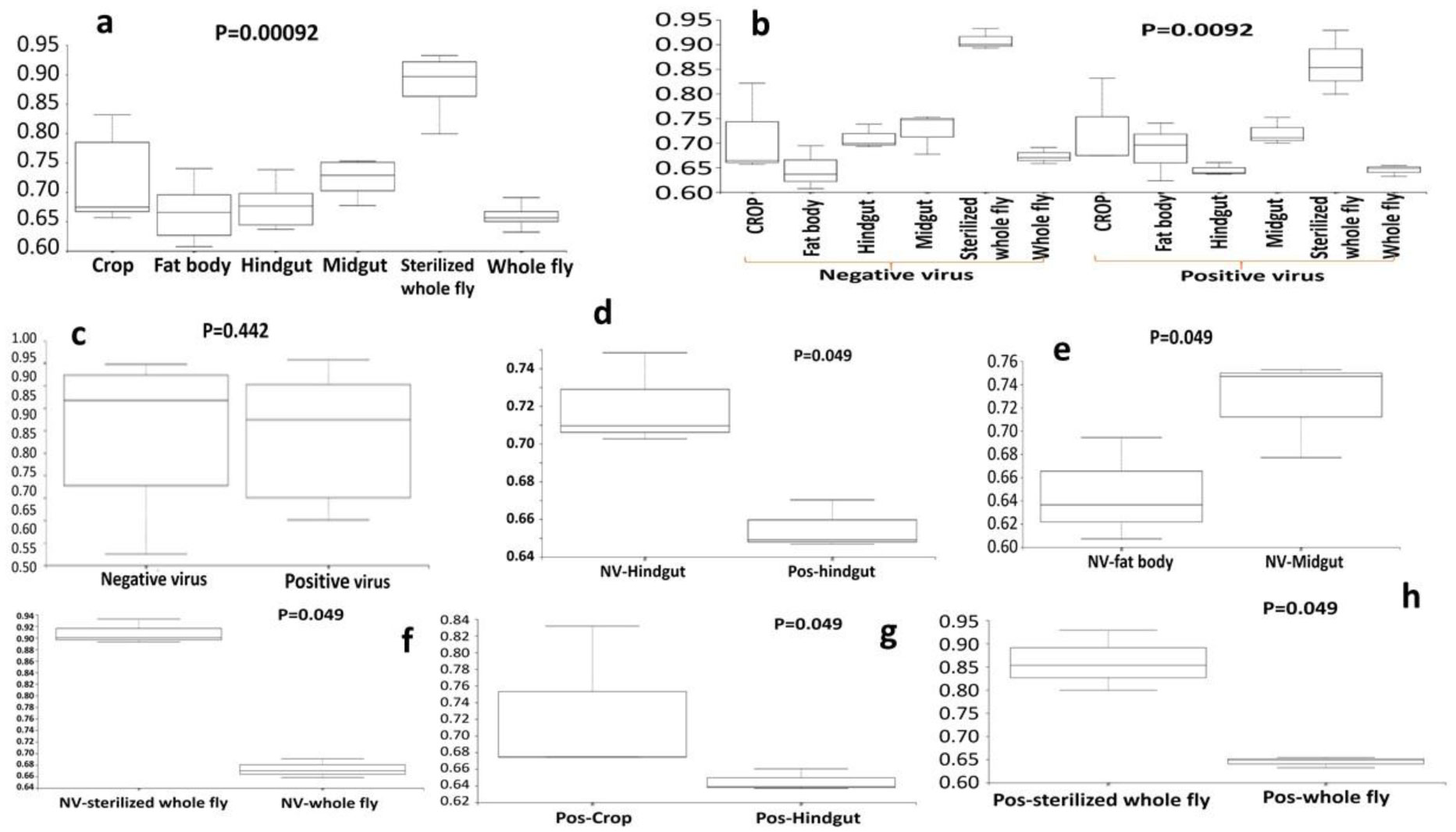
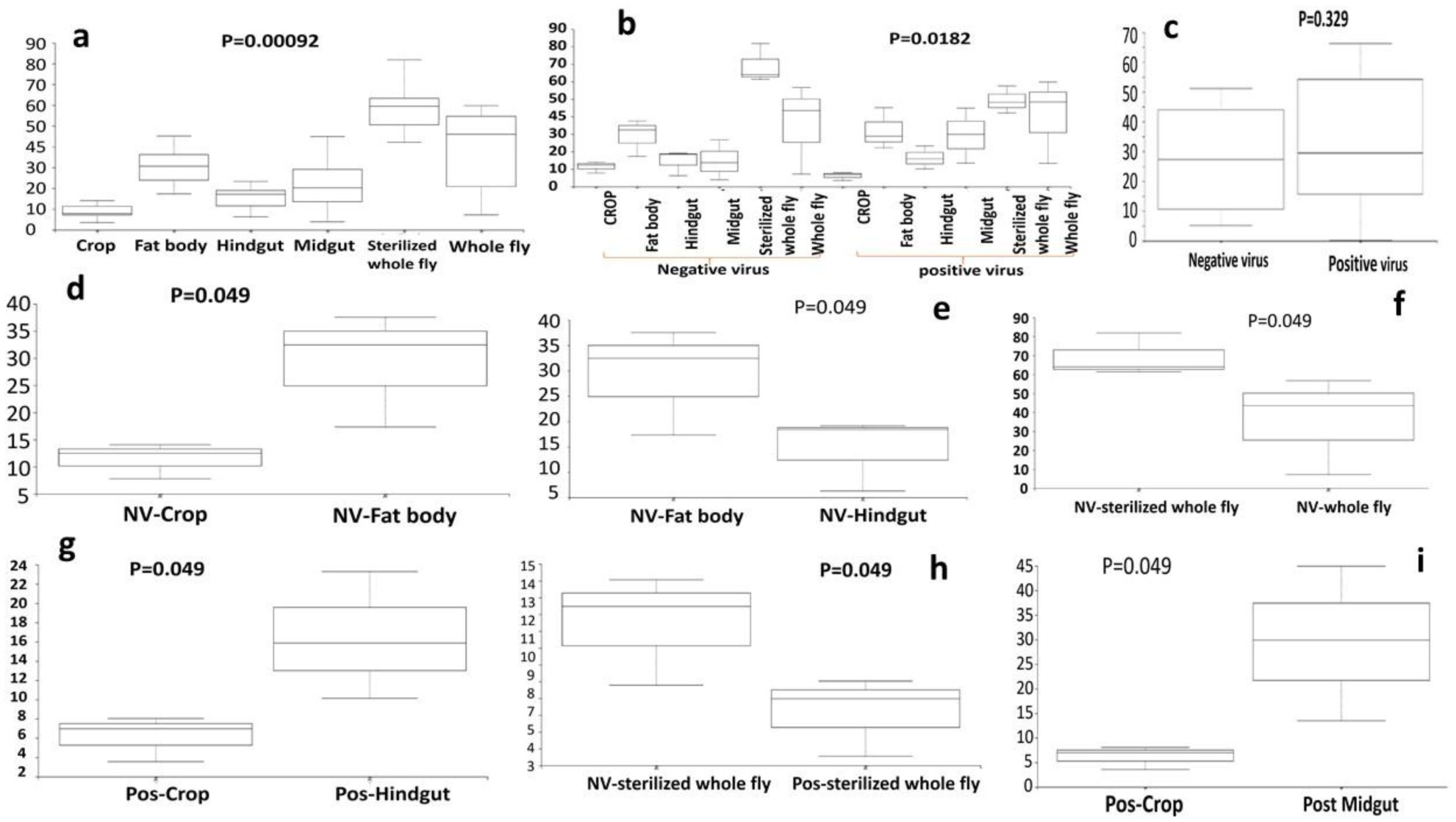
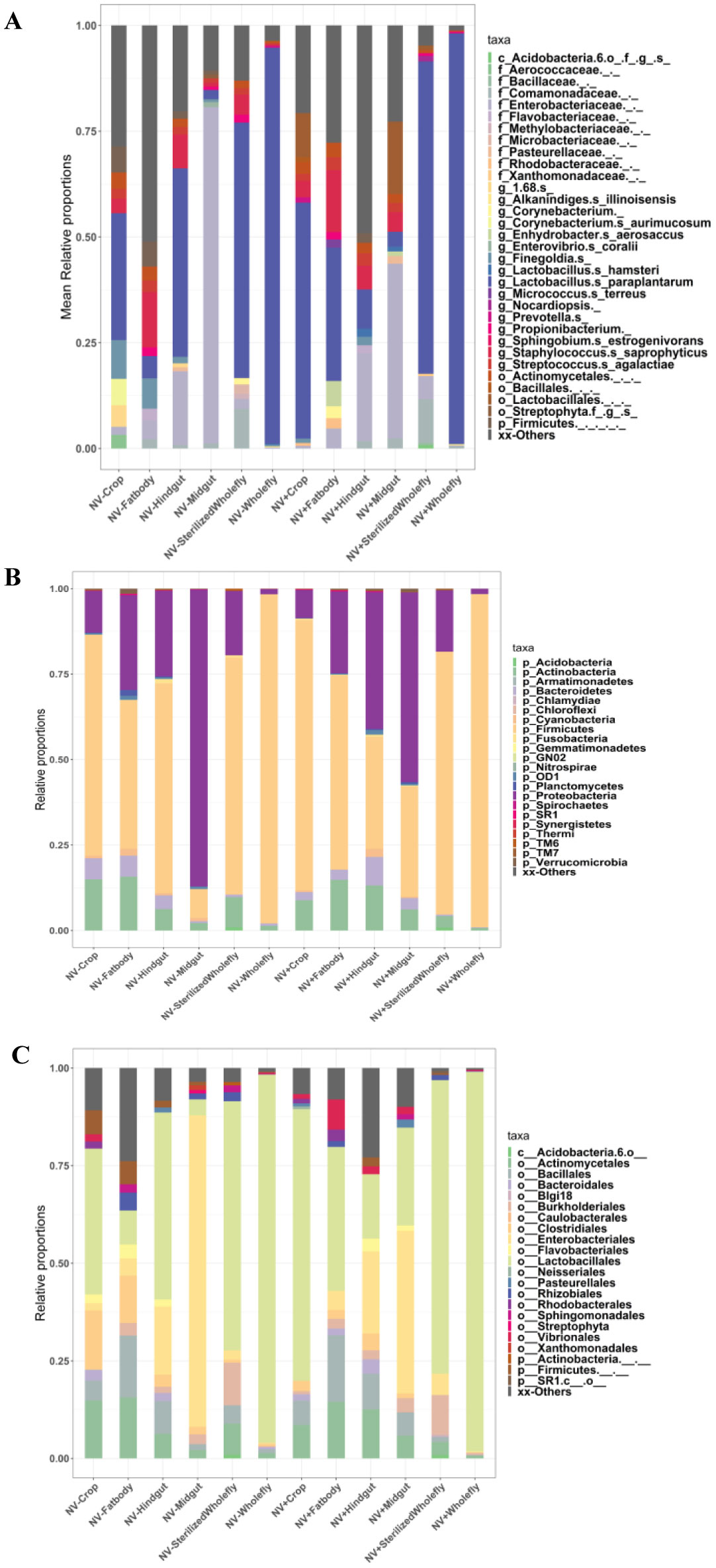
Gastrointestinal microflora is a key component in the maintenance of health and longevity across many species. In humans and mice, nonpathogenic viruses present in the gastrointestinal tract enhance the effects of the native bacterial microbiota. However, it is unclear whether nonpathogenic gastrointestinal viruses, such as Nora virus that infects Drosophila melanogaster, lead to similar observations. Longevity analysis of Nora virus infected (NV+) and uninfected (NV−) D. melanogaster in relationship to presence (B+) or absence (B-) of the native gut bacteria using four different treatment groups, NV+/B+, NV+/B−, NV−/B+, and NV−/B−, was conducted. Data from the longevity results were tested via Kaplan-Meier analysis and demonstrated that Nora virus can be detrimental to the longevity of the organism, whereas bacterial presence is beneficial. These data led to the hypothesis that gastrointestinal bacterial composition varies from NV+ to NV− flies. To test this, NV+ and NV− virgin female flies were collected and aged for 4 days. Surface sterilization followed by dissections of the fat body and the gastrointestinal tract, divided into crop (foregut), midgut, and hindgut, were performed. Ribosomal 16S DNA samples were sequenced to determine the bacterial communities that comprise the microflora in the gastrointestinal tract of NV+ and NV− D. melanogaster. When analyzing operational taxonomic units (OTUs), the data demonstrate that the NV+ samples consist of more OTUs than NV− samples. The NV+ samples were both more rich and diverse in OTUs compared to NV−. When comparing whole body samples to specific organs and organ sections, the whole fly was more diverse in OTUs, whereas the crop was the most rich. These novel data are pertinent in describing where Nora virus infection may be occurring within the gastrointestinal tract, as well as continuing discussion between the relationship of persistent viral and bacterial interaction.
Citation: Makayla Schissel, Rebecca Best, Shelby Liesemeyer, Yuan-De Tan, Darby J. Carlson, Julie J. Shaffer, Nagavardhini Avuthu, Chittibabu Guda, Kimberly A. Carlson. Effect of Nora virus infection on native gut bacterial communities of Drosophila melanogaster[J]. AIMS Microbiology, 2021, 7(2): 216-237. doi: 10.3934/microbiol.2021014
Gastrointestinal microflora is a key component in the maintenance of health and longevity across many species. In humans and mice, nonpathogenic viruses present in the gastrointestinal tract enhance the effects of the native bacterial microbiota. However, it is unclear whether nonpathogenic gastrointestinal viruses, such as Nora virus that infects Drosophila melanogaster, lead to similar observations. Longevity analysis of Nora virus infected (NV+) and uninfected (NV−) D. melanogaster in relationship to presence (B+) or absence (B-) of the native gut bacteria using four different treatment groups, NV+/B+, NV+/B−, NV−/B+, and NV−/B−, was conducted. Data from the longevity results were tested via Kaplan-Meier analysis and demonstrated that Nora virus can be detrimental to the longevity of the organism, whereas bacterial presence is beneficial. These data led to the hypothesis that gastrointestinal bacterial composition varies from NV+ to NV− flies. To test this, NV+ and NV− virgin female flies were collected and aged for 4 days. Surface sterilization followed by dissections of the fat body and the gastrointestinal tract, divided into crop (foregut), midgut, and hindgut, were performed. Ribosomal 16S DNA samples were sequenced to determine the bacterial communities that comprise the microflora in the gastrointestinal tract of NV+ and NV− D. melanogaster. When analyzing operational taxonomic units (OTUs), the data demonstrate that the NV+ samples consist of more OTUs than NV− samples. The NV+ samples were both more rich and diverse in OTUs compared to NV−. When comparing whole body samples to specific organs and organ sections, the whole fly was more diverse in OTUs, whereas the crop was the most rich. These novel data are pertinent in describing where Nora virus infection may be occurring within the gastrointestinal tract, as well as continuing discussion between the relationship of persistent viral and bacterial interaction.

| [1] | Gorbach SL (1996) Microbiology of the gastrointestinal tract. Medical Microbiology Galveston: Universtity of Texax Medical Branch at Galveston. |
| [2] |
Dantoft W, Lundin D, Esfahani SS, et al. (2016) The POU/Oct transcription factor Pdm1/nub is necessary for a beneficial gut microbiota and normal lifespan of Drosophila. J Innate Immun 8: 412-426. doi: 10.1159/000446368

|
| [3] |
Reiter LT, Potocki L, Chien S, et al. (2001) A systematic analysis of human disease-associated gene sequences in Drosophila melanogaster. Genome Res 11: 1114-1125. doi: 10.1101/gr.169101
|
| [4] |
Newell PD, Chaston JM, Wang Y, et al. (2014) In vivo function and comparative genomic analyses of the Drosophila gut microbiota identify candidate symbiosis factors. Front Microbiol 5: 576. doi: 10.3389/fmicb.2014.00576
|
| [5] |
Strigini M, Leulier F (2016) The role of the microbial environment in Drosophila post-embryonic development. Dev Comp Immunol 64: 39-52. doi: 10.1016/j.dci.2016.01.017
|
| [6] |
Sommer AJ, Newell PD (2019) Metabolic basis for mutualism between gut bacteria and its impact on the Drosophila melanogaster host. Appl Environ Micro 85: e01182-18. doi: 10.1128/AEM.01882-18
|
| [7] |
Brummel T, Ching A, Seroude L, et al. (2004) Drosophila lifespan enhancement by exogenous bacteria. Proc Natl Acad Sci 101: 12974-12979. doi: 10.1073/pnas.0405207101
|
| [8] |
Storelli G, Defaye A, Erkosar B, et al. (2011) Lactobacillus plantarum promotes drosophila systemic growth by modulating hormonal signals through TOR-dependent nutrient sensing. Cell Metab 14: 403-414. doi: 10.1016/j.cmet.2011.07.012
|
| [9] |
Nehme NT, Liégeois S, Kele B, et al. (2007) A model of bacterial intestinal infections in Drosophila melanogaster. PLoS Pathog 3: e173. doi: 10.1371/journal.ppat.0030173
|
| [10] |
Fast D, Kostiuk B, Foley E, et al. (2018) Commensal pathogen competition impacts host viability. Proc Natl Acad Sci 115: 7099-7104. doi: 10.1073/pnas.1802165115
|
| [11] |
Wong ACN, Vanhove AS, Watnick PI (2016) The interplay between intestinal bacteria and host metabolism in health and disease: Lessons from Drosophila melanogaster. Dis Model Mech 9: 271-281. doi: 10.1242/dmm.023408
|
| [12] |
Buchon N, Broderick NA, Chakrabarti S, et al. (2009) Invasive and indigenous microbiota impact intestinal stem cell activity through multiple pathways in Drosophila. Genes Dev 23: 2333-2344. doi: 10.1101/gad.1827009
|
| [13] |
Sharon G, Segal D, Ringo JM, et al. (2010) Commensal bacteria play a role in mating preference of Drosophila melanogaster. Proc Natl Acad Sci 107: 20051-20056. doi: 10.1073/pnas.1009906107
|
| [14] |
Xi Z, Ramirez JL, Dimopoulos G (2008) The Aedes aegypti toll pathway controls Dengue virus infection. PLoS Pathog 4: e1000098. doi: 10.1371/journal.ppat.1000098
|
| [15] |
Wu P, Sun P, Nie K, et al. (2019) A gut commensal bacterium promotes mosquito permissiveness to arboviruses. Cell Host Microbe 25: 101-112.e5. doi: 10.1016/j.chom.2018.11.004
|
| [16] |
Barletta ABF, Nascimento-Silva MCL, Talyuli OAC, et al. (2017) Microbiota activates IMD pathway and limits Sindbis infection in Aedes aegypti. Parasit Vectors 10: 103. doi: 10.1186/s13071-017-2040-9
|
| [17] |
Carissimo G, Pondeville E, McFarlane M, et al. (2015) Antiviral immunity of Anopheles gambiae is highly compartmentalized, with distinct roles for RNA interference and gut microbiota. Proc Natl Acad Sci 112: E176-E185. doi: 10.1073/pnas.1412984112
|
| [18] |
Sansone CL, Cohen J, Yasunga A, et al. (2015) Microbiota-dependent priming of antiviral intestinal immunity in Drosophila. Cell Host Microbe 18: 571-581. doi: 10.1016/j.chom.2015.10.010
|
| [19] |
Franchini A, Mandrioli M, Franceschi C, et al. (2012) Morpho-functional changes of fat body in bacteria fed Drosophila melanogaster strains. J Mol Hist 43: 243-251. doi: 10.1007/s10735-011-9382-y
|
| [20] | Shi J, Song Y, Shentu X, et al. (2021) Anitmicrobials affect the fat body microbiome and increase the brown planthopper mortality. Front Microbiol 12: 644897. |
| [21] |
Nelson AM, Elftman MD, Pinto AK, et al. (2013) Murine norovirus infection does not cause major disruptions in the murine intestinal microbiota. Microbiome 1: 17. doi: 10.1186/2049-2618-1-7
|
| [22] |
Sartor RB, Wu GD (2017) Roles for intestinal bacteria, viruses, and fungi in pathogenesis of inflammatory bowel diseases and therapeutic approaches. Gastroenterol 152: 327-339. doi: 10.1053/j.gastro.2016.10.012
|
| [23] | Munn B, Ericson B, Carlson DJ, et al. (2015) Detecting the presence of Nora virus in Drosophila utilizing single fly RT-PCR. Bioscene 41: 40-44. |
| [24] |
Ekström JO, Habayeb MS, Srivastava V, et al. (2011) Drosophila Nora virus capsid proteins differ from those of other picorna-like viruses. Virus Res 160: 51-58. doi: 10.1016/j.virusres.2011.05.006
|
| [25] |
Habayeb MS, Ekengren SK, Hultmark D (2006) Nora virus, a persistent virus in Drosophila, defines a new picorna-like virus family. J Gen Virol 87: 3045-3051. doi: 10.1099/vir.0.81997-0
|
| [26] |
Habayeb MS, Cantera R, Casanova G, et al. (2009) The Drosophila Nora virus is an enteric virus, transmitted via feces. J Invertebr Pathol 101: 29-33. doi: 10.1016/j.jip.2009.02.003
|
| [27] |
Rogers A, Towery L, McCown A, et al. (2020) Impaired geotaxis as a novel phenotype of Nora virus infection of Drosophila melanogaster. Scientifica 2020: Article ID 1804510. doi: 10.1155/2020/1804510
|
| [28] |
Karst SM (2016) The influence of commensal bacteria on infection with enteric viruses. Nat Rev Microbiol 14: 197-204. doi: 10.1038/nrmicro.2015.25
|
| [29] |
Cordes EJ, Licking-Murray KD, Carlson KA (2013) Differential gene expression related to Nora virus infection of Drosophila melanogaster. Virus Res 175: 95-100. doi: 10.1016/j.virusres.2013.03.021
|
| [30] |
Hedengren M, Asling B, Dushay MS, et al. (1999) Relish, a central factor in the control of humoral but not cellular immunity in Drosophila. Mol Cell 4: 827-837. doi: 10.1016/S1097-2765(00)80392-5
|
| [31] |
Caporaso JG, Kuczynski J, Stombaugh J, et al. (2010) QIIME allows analysis of high-throughput community sequencing data. Nat Methods 7: 335-336. doi: 10.1038/nmeth.f.303
|
| [32] |
Callahan BJ, McMurdie PJ, Rosen MJ, et al. (2016) DADA2: High-resolution sample inference from Illumina amplicon data. Nat Methods 13: 581-583. doi: 10.1038/nmeth.3869
|
| [33] |
Bokulich NA, Subramanian S, Faith JJ, et al. (2013) Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat Methods 10: 57-59. doi: 10.1038/nmeth.2276
|
| [34] |
Spellerberg IF, Fedor PJ (2003) A tribute to Claude-Shannon (1916-2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon-Wiener’ Index. Glob Ecol Biogeogr 12: 177-179. doi: 10.1046/j.1466-822X.2003.00015.x
|
| [35] |
Pielou EC (1981) The usefulness of ecological models: A stock-taking. Q Rev Biol 56: 17-31. doi: 10.1086/412081
|
| [36] |
Mulder CPH, Bazeley-White E, Dimitrakopoulos PG, et al. (2004) Species evenness and productivity in experimental plant communities. Oikos 107: 50-63. doi: 10.1111/j.0030-1299.2004.13110.x
|
| [37] |
Faith DP (1992) Conservation evaluation and phylogenetic diversity. Biol Conserv 61: 1-10. doi: 10.1016/0006-3207(92)91201-3
|
| [38] |
Lozupone C, Knight R (2005) UniFrac: A new phylogenetic method for comparing microbial communities. Appl Environ Microbiol 71: 8228-8235. doi: 10.1128/AEM.71.12.8228-8235.2005
|
| [39] | Sultan R, Stampas A, Goldberg MB, et al. (2001) Drug resistance of bacteria commensal with Drosophila melanogaster in laboratory cultures. Dros Inf Serv 84: 175-180. |
| [40] |
Kernbauer E, Ding Y, Cadwell K (2014) An enteric virus can replace the beneficial function of commensal bacteria. Nature 516: 94-98. doi: 10.1038/nature13960
|
| [41] |
Johnson KN, Christian PD (1998) The novel genome organization of the insect picorna-like virus Drosophila C virus suggests this virus belongs to a previously undescribed virus family. J Gen Virol 79: 191-203. doi: 10.1099/0022-1317-79-1-191
|
| [42] | Arnold PA, Johnson KN, White CR (2013) Physiological and metabolic consequences of viral infection in Drosophila melanogaster. J Exp Biol 216: 3350-3357. |
| [43] |
Rozas-Dennis GS, Cazzaniga NJ (2000) Effects of Triatoma virus (TrV) on the fecundity and moulting of Triatoma infestans (Hemiptera: Reduviidae). Ann Trop Med Parasitol 94: 633-641. doi: 10.1080/00034983.2000.11813586
|
| [44] |
Clark RI, Salazar A, Yamada R, et al. (2015) Distinct shifts in microbiota composition during Drosophila aging impair intestinal function and drive mortality. Cell Rep 12: 1656-1667. doi: 10.1016/j.celrep.2015.08.004
|
| [45] |
Petkau K, Parsons BD, Duggal A, et al. (2014) A deregulated intestinal cell cycle program disrupts tissue homeostasis without affecting longevity in Drosophila. J Biol Chem 289: 28719-29. doi: 10.1074/jbc.M114.578708
|
| [46] |
Yamada R, Deshpande SA, Bruce KD, et al. (2015) Microbes promote amino acid harvest to rescue undernutrition in Drosophila. Cell Rep 10: 865-72. doi: 10.1016/j.celrep.2015.01.018
|
| [47] |
Galenza A, Hutchinson J, Campbell SD, et al. (2016) Glucose modulates Drosophila longevity and immunity independent of the microbiota. Biol Open 5: 165-73. doi: 10.1242/bio.015016
|
| [48] | Téfit MA, Leulier F (2017) Lactobacillus plantarum favors the early emergence of fit and fertile adult Drosophila upon chronic undernutrition. J Exp Biol 220: 900-07. |
| [49] |
Obata F, Fons CO, Gould AP (2018) Early-life exposure to low-dose oxidants can increase longevity via microbiome remodelling in Drosophila. Nat Commun 9: 975. doi: 10.1038/s41467-018-03070-w
|
| [50] |
Ren C, Webster P, Finkel SE, et al. (2007) Increased internal and external bacterial load during Drosophila aging without life-span trade-off. Cell Metab 6: 144-152. doi: 10.1016/j.cmet.2007.06.006
|
| [51] |
Matsumoto M, Kurihara S, Kibe R, et al. (2011) Longevity in mice is promoted by probiotic-induced suppression of colonic senescence dependent on upregulation of gut bacterial polyamine production. PLoS One 6: e23652. doi: 10.1371/journal.pone.0023652
|
| [52] |
Erkosar B, Leulier F (2014) Transient adult microbiota, gut homeostasis and longevity: Novel insights from the Drosophila model. FEBS Letters 588: 4250-4257. doi: 10.1016/j.febslet.2014.06.041
|
| [53] |
Han G, Lee HJ, Jeong SE, et al. (2017) Comparative analysis of Drosophila melanogaster gut microbiota with respect to host strain, sex, and age. Microb Ecol 74: 207-216. doi: 10.1007/s00248-016-0925-3
|
| [54] |
Grandison RC, Wong R, Bass TM, et al. (2009) Effect of standardized dietary restriction protocol on multiple laboratory strains of Drosophila melanogaster. PLoS One 4: e4067. doi: 10.1371/journal.pone.0004067
|
| [55] |
Lee HY, Lee SH, Lee JH, et al. (2019) The role of commensal microbes in the lifespan of Drosophila melanogaster. Aging 11: 4611-4640. doi: 10.18632/aging.102073
|
| [56] |
Wong CN, Ng P, Douglas AE (2011) Low-diversity bacterial community in the gut of the fruitfly. Env Microbiol 13: 1889-1900. doi: 10.1111/j.1462-2920.2011.02511.x
|
| [57] |
Dang AT, Cotton S, Sankaran-Walters S, et al. (2012) Evidence of an increased pathogenic footprint in the lingual microbiome of untreated HIV infected patients. BMC Microbiol 12: 1-10. doi: 10.1186/1471-2180-12-1
|
| [58] |
Manna S, Baindara P, Mandal SM (2020) Molecular pathogenesis of secondary bacterial infection associated to viral infections including SARS-CoV-2. J Infect Public Health 13: 1397-1404. doi: 10.1016/j.jiph.2020.07.003
|
| [59] | Smith H, Sweet C (2002) Cooperation between viral and bacterial pathogens in causing human respiratory disease. Polymicrobial Diseases Washington: ASM Press. |
| [60] |
Malacrinò A, Campolo O, Medina RF, et al. (2018) Instar- and host-associated differentiation of bacterial communities in the Mediterranean fruit fly Ceratitis capitata. PLoS One 13: e0194131. doi: 10.1371/journal.pone.0194131
|
| [61] |
Gontang EA, Aylward FO, Carlos C, et al. (2017) Major changes in microbial diversity and community composition across gut sections of a juvenile Panchlora cockroach. PLoS One 12: e0177189. doi: 10.1371/journal.pone.0177189
|
| [62] |
Miguel-Aliaga I, Jasper H, Lemaitre B (2018) Anatomy and physiology of the digestive tract of Drosophila melanogaster. Genet 210: 357-396. doi: 10.1534/genetics.118.300224
|
| [63] |
Cohen E, Sawyer JK, Peterson NG, et al. (2020) Physiology, development, and disease modeling in the Drosophila excretory system. Genet 214: 235-264. doi: 10.1534/genetics.119.302289
|
| [64] | Zhang Y, Yongmei X (2014) Fat body development and its function in energy storage and nutrient sensing in Drosophila melanogaster. J Tissue Sci Eng 6: 1. |
| [65] |
Chandler JA, Lang J, Bhatnagar S, et al. (2011) Bacterial communities of diverse Drosophila species: Ecological context of a host-microbe model system. PLoS Genet 7: e1002272. doi: 10.1371/journal.pgen.1002272
|
| [66] |
Jones RM, Luo L, Ardita CS, et al. (2013) Symbiotic lactobacilli stimulate gut epithelial proliferation via Nox-mediated generation of reactive oxygen species. EMBO J 32: 3017-3028. doi: 10.1038/emboj.2013.224
|
| [67] |
Saxena D, Li Y, Yang L, et al. (2012) Human microbiome and HIV/AIDS. Curr HIV/AIDS Rep 9: 44-51. doi: 10.1007/s11904-011-0103-7
|
| [68] |
Jones MK, Watanabe M, Zhu S, et al. (2014) Enteric bacteria promote human and mouse norovirus infection of B Cells. Science 346: 755-759. doi: 10.1126/science.1257147
|
 microbiol-07-02-014-s001.pdf microbiol-07-02-014-s001.pdf |
 |

Figures(7)

Makayla Schissel, Rebecca Best, Shelby Liesemeyer, Yuan-De Tan, Darby J. Carlson, Julie J. Shaffer, Nagavardhini Avuthu, Chittibabu Guda, Kimberly A. Carlson. Effect of Nora virus infection on native gut bacterial communities of Drosophila melanogaster[J]. AIMS Microbiology, 2021, 7(2): 216-237. doi: 10.3934/microbiol.2021014







 DownLoad:
DownLoad: