Citation: Hamed Azizollahi, Marion Darbas, Mohamadou M. Diallo, Abdellatif El Badia, Stephanie Lohrengel. EEG in neonates: Forward modeling and sensitivity analysis with respect to variations of the conductivity[J]. Mathematical Biosciences and Engineering, 2018, 15(4): 905-932. doi: 10.3934/mbe.2018041

| [1] | [ Z. Akalin Acar,S. Makeig, Effects of Forward Model Errors on EEG Source Localization, Brain Topogrography, 26 (2013): 378-396. |
| [2] | [ A. Alonso-Rodriguez,J. Camano,R. Rodriguez,A. Valli, Assessment of two approximation methods for the inverse problem of electroencephalography, Int. J. of Numerical Analysis and Modeling, 13 (2016): 587-609. |
| [3] | [ H. Azizollahi,A. Aarabi,F. Wallois, Effects of uncertainty in head tissue conductivity and complexity on EEG forward modeling in neonates, Hum. Brain Ma, 37 (2016): 3604-3622. |
| [4] | [ H. T. Banks,D. Rubio,N. Saintier, Optimal design for parameter estimation in EEG problems in a 3D multilayered domain, Mathematical Biosciences and Engineering, 12 (2015): 739-760. |
| [5] | [ M. Bauer,S. Pursiainen,J. Vorwerk,H. Köstler,C. H. Wolters, Comparison Study for Whitney (Raviart-Thomas)-Type Source Models in Finite-Element-Method-Based EEG Forward Modeling, IEEE Trans. Biomed. Eng., 62 (2015): 2648-2656. |
| [6] | [ J. Borggaard and V. L. Nunes, Fréchet Sensitivity Analysis for Partial Differential Equations with Distributed Parameters, American Control Conference, San Francisco, 2011. |
| [7] | [ H. Brezis, Functional Analysis, Sobolev Spaces And Partial Differential Equations, Universitext. Springer, New York, 2011. |
| [8] | [ P. G. Ciarlet, The Finite Element Method for Elliptic Problems, North Holland, New York, 1978. |
| [9] | [ M. Clerc,J. Kybic, Cortical mapping by Laplace-Cauchy transmission using a boundary element method, Journal on Inverse Problems, 23 (2007): 2589-2601. |
| [10] | [ M. Clerc, J. Leblond, J. -P. Marmorat and T. Papadopoulo, Source localization using rational approximation on plane sections, Inverse Problems, 28 (2012), 055018, 24 pp. |
| [11] | [ M. Darbas, M. M. Diallo, A. El Badia and S. Lohrengel, An inverse dipole source problem in inhomogeneous media: application to the EEG source localization in neonates, in preparation. |
| [12] | [ A. El Badia,T. Ha Duong, An inverse source problem in potential analysis, Inverse Problems, 16 (2000): 651-663. |
| [13] | [ A. El Badia,M. Farah, Identification of dipole sources in an elliptic equation from boundary measurements, J. Inv. Ill-Posed Problems, 14 (2006): 331-353. |
| [14] | [ A. El Badia and M. Farah, A stable recovering of dipole sources from partial boundary measurements, Inverse Problems, 26 (2010), 115006, 24pp. |
| [15] | [ Q. Fang and D. A. Boas, Tetrahedral mesh generation from volumetric binary and grayscale images, EEE International Symposium on Biomedical Imaging: From Nano to Macro, (2009), SBI? 09. Boston, Massachusetts, USA, 1142–1145. |
| [16] | [ M. Farah, Problémes Inverses de Sources et Lien avec l'Electro-encéphalo-graphie, Thése de doctorat, Université de Technologie de Compiégne, 2007. |
| [17] | [ O. Faugeras, F. Clément, R. Deriche, R. Keriven, T. Papadopoulo, J. Roberts, T. Viéville, F. Devernay, J. Gomes, G. Hermosillo, P. Kornprobst and D. Lingrand, The Inverse EEG and MEG Problems: The Adjoint State Approach I: The Continuous Case ,Rapport de recherche, 1999. |
| [18] | [ P. Gargiulo,P. Belfiore,E. A. Friogeirsson,S. Vanhalato,C. Ramon, The effect of fontanel on scalp EEG potentials in the neonate, Clin. Neurophysiol, 126 (2015): 1703-1710. |
| [19] | [ D. Gilbarg and N. S. Trudinger, Elliptic Partial Differential Equations of Second Order Springer, Berlin, 1977. |
| [20] | [ R. Grech, T. Cassar, J. Muscat, K. P. Camilleri, S. G. Fabri, M. Zervakis, P. Xanthopoulos, V. Sakkalis and B. Vanrumste, Review on solving the inverse problem in EEG source analysis J. NeuorEng. Rehabil. , 5 (2008). |
| [21] | [ H. Hallez, B. Vanrumste, R. Grech, J. Muscat, W. De Clercq, A. Vergult, Y. d'Asseler, K. P. Camilleri, S. G. Fabri, S. Van Huffel and I. Lemahieu, Review on solving the forward problem in EEG source analysis J. NeuorEng. Rehabil., 4 (2007). |
| [22] | [ M. Hämäläinen,R. Hari,J. Ilmoniemi,J. Knuutila,O. V. Lounasmaa, Magnetoencephalography-theory, instrumentation, and applications to noninvasive studies of the working human brain, Rev. Mod. Phys., 65 (1993): 413-497. |
| [23] | [ F. Hecht, O. Pironneau, A. Le Hyaric and K. Ohtsuka, FreeFem++ Manual, 2014. |
| [24] | [ E. Hernández,R. Rodríguez, Finite element Approximation of Spectral Problems with Neumann Boundary Conditions on curved domains, Math. Comp., 72 (2002): 1099-1115. |
| [25] | [ R. Kress, Linear Integral Equations, Second edition, Applied Mathematical Sciences 82, Spinger-Verlag, 1999. |
| [26] | [ J. Kybic,M. Clerc,T. Abboud,O. Faugeras,R. Keriven,T. Papadopoulo, A common formalism for the integral formulations of the forward EEG Problem, IEEE Transactions on Medical Imaging, 24 (2005): 12-28. |
| [27] | [ J. Kybic,M. Clerc,O. Faugeras,R. Keriven,T. Papadopoulo, Fast multipole acceleration of the MEG/EEG boundary element method, Physics in Medicine and Biology, 50 (2005): 4695-4710. |
| [28] | [ J. Kybic,M. Clerc,T. Abboud,O. Faugeras,R. Keriven,T. Papadopoulo, Generalized head models for MEG/EEG: Boundary element method beyond nested volumes, Phys. Med. Biol., 51 (2006): 1333-1346. |
| [29] | [ J. Leblond, Identifiability properties for inverse problems in EEG data processing and medical engineering, with observability and optimization issues, Acta Applicandae Mathematicae, 135 (2015): 175-190. |
| [30] | [ S. Lew,D. D. Silva,M. Choe,P. Ellen Grant,Y. Okada,C. H. Wolters,M. S. Hämäläinen, Effects of sutures and fontanels on MEG and EEG source analysis in a realistic infant head model, Neuroimage, 76 (2013): 282-293. |
| [31] | [ T. Medani,D. Lautru,D. Schwartz,Z. Ren,G. Sou, FEM method for the EEG forward problem and improvement based on modification of the saint venant's method, Progress In Electromagnetic Research, 153 (2015): 11-22. |
| [32] | [ J. C. de Munck,M. J. Peters, A fast method to compute the potential in the multisphere model, IEEE Trans. Biomed. Eng., 40 (1993): 1166-1174. |
| [33] | [ Odabaee,A. Tokariev,S. Layeghy,M. Mesbah,P. B. Colditz,C. Ramon,S. Vanhatalo, Neonatal EEG at scalp is focal and implies high skull conductivity in realistic neonatal head models, NeuroImage, 96 (2014): 73-80. |
| [34] | [ P. A. Raviart and J. M. Thomas, Introduction á l'Analyse Numérique des Equations aux Dérivées Partielles, Masson, Paris, 1983. |
| [35] | [ N. Roche-Labarbe,A. Aarabi,G. Kongolo,C. Gondry-Jouet,M. Dümpelmann,R. Grebe,F. Wallois, High-resolution electroencephalography and source localization in neonates, Human Brain Mapping, 29 (2008): 167-76. |
| [36] | [ C. Rorden,L. Bonilha,J. Fridriksson,B. Bender,H. O. Karnath, Age-specific CT and MRI templates for spatial normalization, NeuroImage, 61 (2012): 957-965. |
| [37] | [ M. Schneider, A multistage process for computing virtual dipole sources of EEG discharges from surface information, IEEE Trans. on Biomed. Eng., 19, 1-19. |
| [38] | [ M. I. Troparevsky, D. Rubio and N. Saintier, Sensitivity analysis for the EEG forward problem Frontiers in Computational Neuroscience, 4 (2010), p138. |
| [39] | [ J. Vorwerk,J. H. Cho,S. Rampp,H. Hamer,T. T. Knösche,C. H. Wolters, A guideline for head volume conductor modeling in EEG and MEG, NeuroImage, 100 (2014): 590-607. |
| [40] | [ C. H. Wolters,H. Köstler,C. Möller,J. Härdtlein,L. Grasedyck,W. Hackbusch, Numerical mathematics of the subtraction approach for the modeling of a current dipole in EEG source reconstruction using finite element head models, SIAM J. Sci. Comput., 30 (2007): 24-45. |
| [41] | [ C. H. Wolters,H. Köstler,C. Möller,J. Härdtlein,A. Anwander, Numerical approaches for dipole modeling in finite element method based source analysis, Int. Congress Ser., 1300 (2007): 189-192. |
| [42] | [ Z. Zhang, A fast method to compute surface potentials generated by dipoles within multilayer anisotropic spheres, Phys. Med. Biol., 40 (1995): 335-349. |
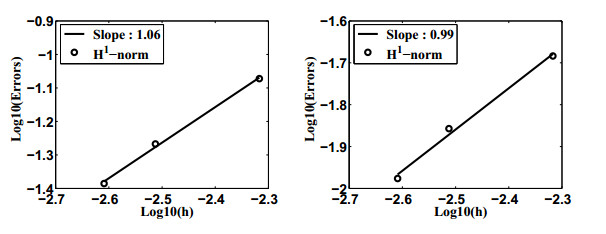
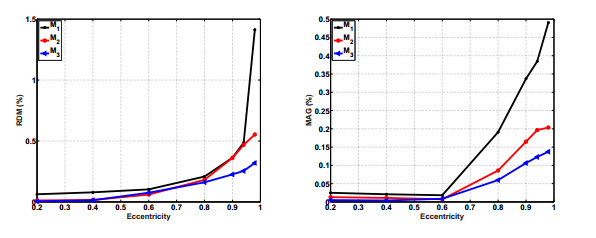
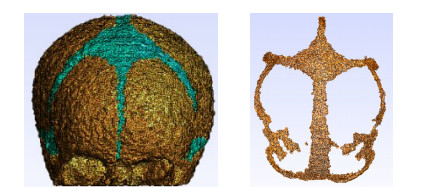
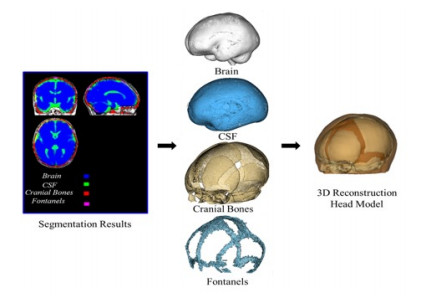

Figures(12) / Tables(2)

Hamed Azizollahi, Marion Darbas, Mohamadou M. Diallo, Abdellatif El Badia, Stephanie Lohrengel. EEG in neonates: Forward modeling and sensitivity analysis with respect to variations of the conductivity[J]. Mathematical Biosciences and Engineering, 2018, 15(4): 905-932. doi: 10.3934/mbe.2018041







 DownLoad:
DownLoad: