Hiking is one of the most popular leisure sport activities practiced in the Alps during the summer season, but bears the risk of mountain emergencies, accidents, and fatalities. This paper provides an updated analysis of hiking accidents for the years 2015 to 2021 in the Austrian Alps, thereby outlining fatal and non-fatal accident characteristics.
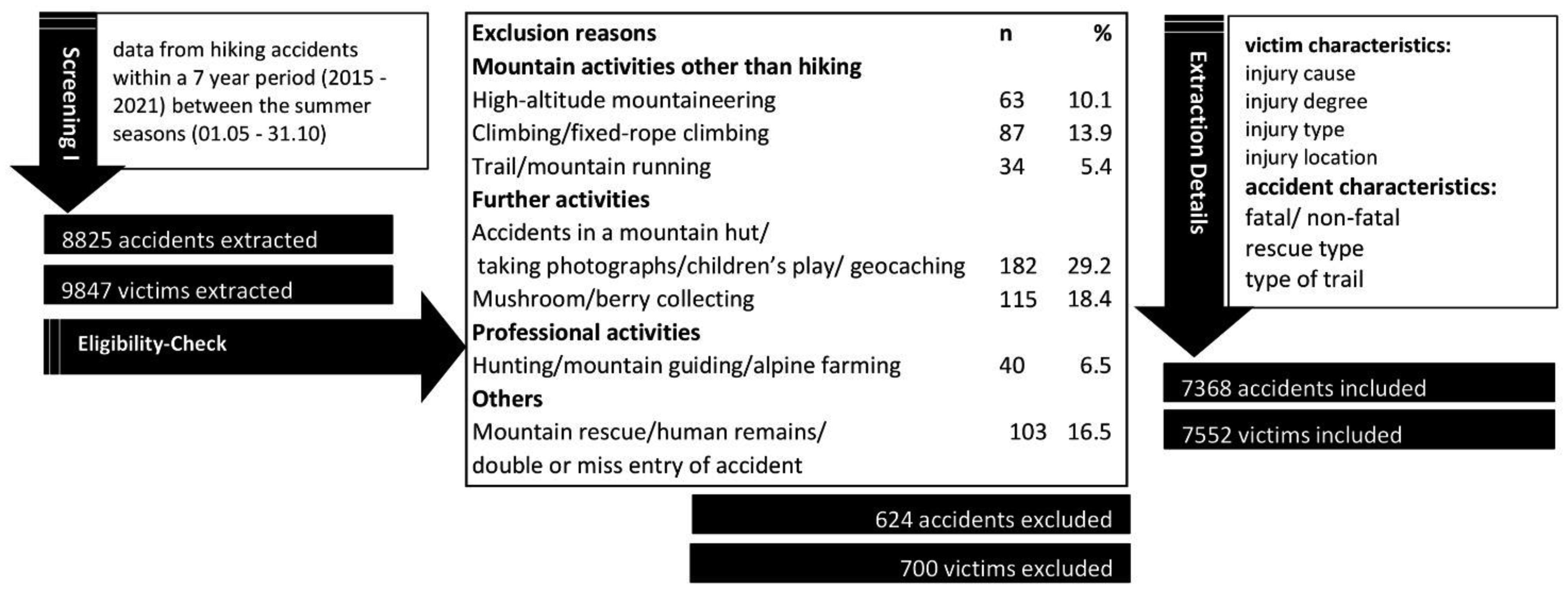
For this retrospective analysis, mountain hiking accidents documented by the Austrian Alpine Police during a 7-year period were screened for potential exclusion criteria. The final sample size consisted of 7368 accidents and 7552 victims. The outcome measures were mainly specified by sex, age, injury degree, injury location, pathophysiological characteristics, and cause of injury.
The overall annual number of accidents showed a continuous increase from 428 in 2015 to 544 in 2021. In total, 7.1% of the total victims died during the 7-year period, with male hikers being significantly more affected than female hikers (m: 80.8%, f: 19.2%; p ≤ 0.001). The sex specific distribution for non-fatal hiking accidents was 55.9% in women and 44.1% in men. Male victims showed significantly more frequent cardiovascular events (m: 78.5%, f: 21.5%), multiple injuries (m: 60.2%, f: 39.8%), and wounds/blood loss (m: 57.4%, f: 42.6%) than female victims, whereas women showed more fractures (m: 31.5%, f: 68.5%) than men (p ≤ 0.001). Additionally, men were more likely to injure their abdomen/chest (3.7%), head (14.1%), and multiple body parts (26.5%), whereas women were more likely to injure their ankle or foot (42.3%). Finally, men were more likely to have an accident during the ascent (24.1%), whereas women during the descent (69.0%) (p ≤ 0.001).
This paper provides the latest data and a deeper insight into sex-specific characteristics of mountain hiking accidents in the Austrian Alps.
Citation: Linda Rausch, Mirjam Limmer, Elena Pocecco, Gerhard Ruedl, Markus Posch, Martin Faulhaber. Sex-specific analysis of hiking accidents in the Austrian Alps: a follow-up from 2015 to 2021[J]. AIMS Public Health, 2024, 11(1): 160-175. doi: 10.3934/publichealth.2024008
Hiking is one of the most popular leisure sport activities practiced in the Alps during the summer season, but bears the risk of mountain emergencies, accidents, and fatalities. This paper provides an updated analysis of hiking accidents for the years 2015 to 2021 in the Austrian Alps, thereby outlining fatal and non-fatal accident characteristics.
For this retrospective analysis, mountain hiking accidents documented by the Austrian Alpine Police during a 7-year period were screened for potential exclusion criteria. The final sample size consisted of 7368 accidents and 7552 victims. The outcome measures were mainly specified by sex, age, injury degree, injury location, pathophysiological characteristics, and cause of injury.
The overall annual number of accidents showed a continuous increase from 428 in 2015 to 544 in 2021. In total, 7.1% of the total victims died during the 7-year period, with male hikers being significantly more affected than female hikers (m: 80.8%, f: 19.2%; p ≤ 0.001). The sex specific distribution for non-fatal hiking accidents was 55.9% in women and 44.1% in men. Male victims showed significantly more frequent cardiovascular events (m: 78.5%, f: 21.5%), multiple injuries (m: 60.2%, f: 39.8%), and wounds/blood loss (m: 57.4%, f: 42.6%) than female victims, whereas women showed more fractures (m: 31.5%, f: 68.5%) than men (p ≤ 0.001). Additionally, men were more likely to injure their abdomen/chest (3.7%), head (14.1%), and multiple body parts (26.5%), whereas women were more likely to injure their ankle or foot (42.3%). Finally, men were more likely to have an accident during the ascent (24.1%), whereas women during the descent (69.0%) (p ≤ 0.001).
This paper provides the latest data and a deeper insight into sex-specific characteristics of mountain hiking accidents in the Austrian Alps.

| [1] | Richins H, Johnsen S, Hull JS, et al. (2016) Overview of mountain tourism: substantive nature, historical context, areas of focus. Mountain tourism: Experiences, communities, environments and sustainable futures . Wallingford UK: CABI 1-12. https://doi.org/10.1079/9781780644608.0001 |
| [2] | Romeo R, Russo L, Parisi F, et al. Mountain tourism–Towards a more sustainable path (2021). |
| [3] |
Burtscher M, Bachmann O, Hatzl T, et al. (2001) Cardiopulmonary and metabolic responses in healthy elderly humans during a 1-week hiking programme at high altitude. Eur J Appl Physiol 84: 379-386. https://doi.org/10.1007/s004210100393 
|
| [4] | Austrian Alpine AssociationTyrol Declaration to best practice in mountain Sports (2002). Available from: https://theuiaa.org/documents/declarations/UIAA_Declaration_TyrolDeclaration.pdf |
| [5] |
Zürcher S, Jackowski C, Schön CA (2020) Circumstances and causes of death of hikers at different altitudes: A retrospective analysis of hiking fatalities from 2003–2018. Forensic Sci Int 310: 110252. https://doi.org/10.1016/j.forsciint.2020.110252
|
| [6] |
Faulhaber M, Ruedl G, Schneider F, et al. (2020) Characteristics of victims of fall-related accidents during mountain hiking. Int J Environ Res Public Health 17: 1115. https://doi.org/10.3390/ijerph17031115
|
| [7] |
Faulhaber M, Pocecco E, Niedermeier M, et al. (2017) Fall-related accidents among hikers in the Austrian Alps: a 9-year retrospective study. BMJ Open Sport Exerc Med 3: e000304. https://doi.org/10.1136/bmjsem-2017-000304
|
| [8] |
Curran-Sills GM, Karahalios A (2015) Epidemiological trends in search and rescue incidents documented by the Alpine Club of Canada from 1970 to 2005. Wilderness Environ Med 26: 536-543. https://doi.org/10.1016/j.wem.2015.07.001
|
| [9] |
Van Mechelen W, Hlobil H, Kemper HCG (1992) Incidence, severity, aetiology and prevention of sports injuries: a review of concepts. Sports Med 14: 82-99. https://doi.org/10.2165/00007256-199214020-00002
|
| [10] | WHOCardiovascular diseases (CVDs) (2021). Available from: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) |
| [11] |
Rossi L, Behme N, Breuer C (2021) Physical activity of children and adolescents during the COVID-19 pandemic—A scoping review. Int J Environ Res Public Health 18: 11440. https://doi.org/10.3390/ijerph182111440
|
| [12] |
Faulhaber M, Flatz M, Gatterer H, et al. (2007) Prevalence of cardiovascular diseases among alpine skiers and hikers in the Austrian Alps. High Alt Med Biol 8: 245-252. https://doi.org/10.1089/ham.2007.1005
|
| [13] | Österreichischer Alpenverein Anzahl der Mitglieder des Österreichischen Alpenvereins von 2011 bis 2021 (2022). Available from: https://de.statista.com/statistik/daten/studie/1198438/umfrage/mitglieder-des-oesterreichischen-alpenvereins |
| [14] | Deutscher Olympischer SportbundMitgliederzahl des Deutschen Alpenvereins von 2002 bis 2022 (2022). Available from: https://de.statista.com/statistik/daten/studie/215943/umfrage/mitgliederzahl-des-deutschen-alpenvereins |
| [15] |
Pocecco E, Wafa H, Burtscher J, et al. (2022) Mortality in recreational mountain-biking in the Austrian Alps: a retrospective study over 16 years. Int J Environ Res Public Health 19: 11965. https://doi.org/10.3390/ijerph191911965
|
| [16] |
Bigdon SF, Hecht V, Fairhurst PG, et al. (2022) Injuries in alpine summer sports - types, frequency and prevention: a systematic review. BMC Sports Sci Med Rehabil 14: 79. https://doi.org/10.1186/s13102-022-00468-4
|
| [17] | German Alpine ClubGender statistics in the German Alpine Club (2022). Available from: https://www.alpenverein.de/files/dav-geschlechterstatistik-2022.pdf |
| [18] |
Ruedl G, Webhofer M, Linortner I, et al. (2011) ACL injury mechanisms and related factors in male and female carving skiers: a retrospective study. Int J Sports Med 32: 801-806. https://doi.org/10.1055/s-0031-1279719
|
| [19] |
Laapotti S, Keskinen E (2004) Has the difference in accident patterns between male and female drivers changed between 1984 and 2000?. Accid Anal Prev 36: 577-584. https://doi.org/10.1016/S0001-4575(03)00064-2
|
| [20] |
Ambrose AF, Cruz L, Paul G (2015) Falls and fractures: a systematic approach to screening and prevention. Maturitas 82: 85-93. https://doi.org/10.1016/j.maturitas.2015.06.035
|
| [21] | Frohlick S The “hypermasculine” landscape of high-altitude mountaineering (2000). |
| [22] |
Krokosz D, Lipowski M (2022) “No Risk No Fun?”: Determinants of Satisfaction with Life in People Who Engage in Extreme and High-Risk Sports. Int J Environ Res Public Health 19: 13328. https://doi.org/10.3390/ijerph192013328
|
| [23] |
Apollo M, Mostowska J, Legut A, et al. (2023) Gender differences in competitive adventure sports tourism. J Outdoor Rec Tour 42: 100604. https://doi.org/10.1016/j.jort.2022.100604
|
| [24] |
Gasser B (2019) The older the hiker, the more severe the injury-a retrospective analysis of mountain hiking accidents in the Swiss Alps from 2009 to 2018. Praxis 108: 1065-1071. https://doi.org/10.1024/1661-8157/a003328
|


Figures(3) / Tables(5)

Linda Rausch, Mirjam Limmer, Elena Pocecco, Gerhard Ruedl, Markus Posch, Martin Faulhaber. Sex-specific analysis of hiking accidents in the Austrian Alps: a follow-up from 2015 to 2021[J]. AIMS Public Health, 2024, 11(1): 160-175. doi: 10.3934/publichealth.2024008







 DownLoad:
DownLoad: