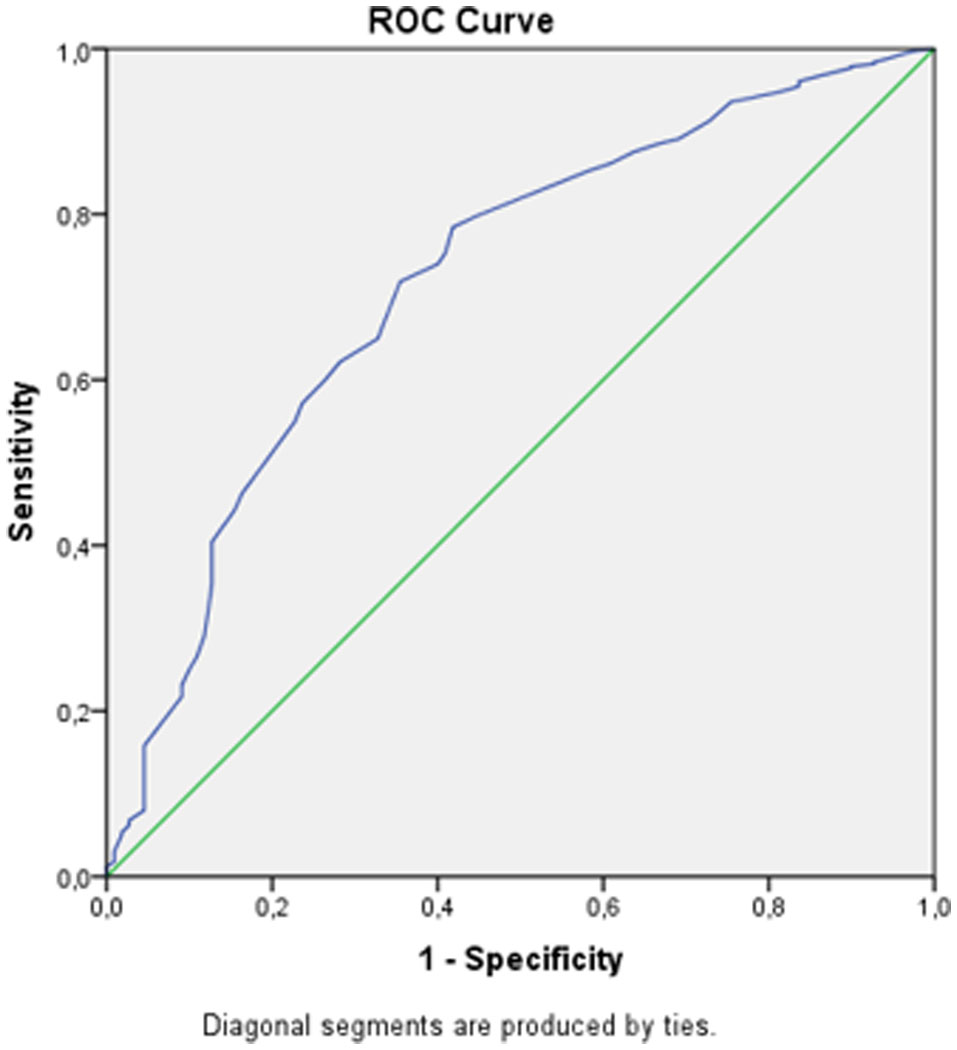
Severe acute malnutrition (SAM) remains a main cause of mortality among children under five years old. Vietnam needs further study to establish the optimal mid-upper-arm circumference (MUAC) cutoff for improving the accuracy of the MUAC indicator in screening SAM children aged 6–59 months. A survey was conducted at all 16 subdistricts across four provinces in Northern Midlands and mountainous areas. The data of 4,764 children showed that an optimal MUAC cutoff of 13.5 cm would allow the inclusion of 65% of children with weight-for-height z-scores (WHZs) below −3SD. A combination of MUAC and WHZ may achieve a higher impact on therapeutic feeding programs for SAM children. The MUAC cutoff of 13.5 cm (65% sensitivity and 72% specificity) should be used as the cutoff for improving and/or preventing SAM status among children under 5 in the Midlands and mountainous areas in Vietnam.
Citation: Tran Thi Hai, Saptawati Bardosono, Luh Ade Ari Wiradnyani, Le Thi Hop, Hoang T. Duc Ngan, Huynh Nam Phuong. The optimal mid-upper-arm circumference cutoffs to screen severe acute malnutrition in Vietnamese children[J]. AIMS Public Health, 2020, 7(1): 188-196. doi: 10.3934/publichealth.2020016
Severe acute malnutrition (SAM) remains a main cause of mortality among children under five years old. Vietnam needs further study to establish the optimal mid-upper-arm circumference (MUAC) cutoff for improving the accuracy of the MUAC indicator in screening SAM children aged 6–59 months. A survey was conducted at all 16 subdistricts across four provinces in Northern Midlands and mountainous areas. The data of 4,764 children showed that an optimal MUAC cutoff of 13.5 cm would allow the inclusion of 65% of children with weight-for-height z-scores (WHZs) below −3SD. A combination of MUAC and WHZ may achieve a higher impact on therapeutic feeding programs for SAM children. The MUAC cutoff of 13.5 cm (65% sensitivity and 72% specificity) should be used as the cutoff for improving and/or preventing SAM status among children under 5 in the Midlands and mountainous areas in Vietnam.

| [1] |
Black R, Victora C, Walker S, et al. (2013) Maternal and child undernutrition and overweight in low-income and middle-income countries. Lancet 382: 427-451. doi: 10.1016/S0140-6736(13)60937-X

|
| [2] |
Black R, Allen L, Bhutta Z, et al. (2008) Maternal and child undernutrition: global and regional exposures and health consequences. Lancet 371: 243-260. doi: 10.1016/S0140-6736(07)61690-0
|
| [3] |
Bhutta Z, Das J, Rizvi A, et al. (2013) Evidence-based interventions for improvement of maternal and child nutrition: What can be done and at what cost? Lancet 382: 452-477. doi: 10.1016/S0140-6736(13)60996-4
|
| [4] | World Health Organization (2003) Guideline: Update on the Management of Severe Acute Malnutrition in Infants and Children. |
| [5] |
Collins S, Sadler K, Dent N, et al. (2006) Key issues in the success of community-based management of severe malnutrition. Food Nutr Bull 27: S49-S82. doi: 10.1177/15648265060273S304
|
| [6] | Huynh P, Tran TN, Roger M, et al. (2014) Development and implementation of a locally produced ready-to-use therapeutic food (RUTF) in Vietnam? Food and Nutrition Bulletin 35: 52-56. |
| [7] | (2011) National Institute of Nutrition and UNICEFInterim Guidelines on the Integrated Management of Acute Malnutrition in Viet Nam. UNICEF. Available from: http://www.unscn.org/files/Announcements/vacancies/UNICEF_Nutrition_Specialist_Consultancy_Notice_Vietnam_May_2014_CSD_2014-01.pdf |
| [8] |
Grellety E, Golden M (2018) Severely malnourished children with a low weight-for-height have similar mortality to those with a low mid-upper-arm-circumference: II. Systematic literature review and meta-analysis. Nutr J 17: 80. doi: 10.1186/s12937-018-0383-5
|
| [9] |
Whye Lian C, Wan Muda WAM, Mohd Hussin ZA, et al. (2012) Factors associated with undernutrition among children in a rural district of Kelantan, Malaysia. Asia Pac J Public Health 24: 330-342. doi: 10.1177/1010539510380737
|
| [10] |
Fernández M, Delchevalerie P, Van Herp M (2010) Accuracy of MUAC in the detection of severe wasting with the new WHO growth standards. Pediatrics 126: e195-e201. doi: 10.1542/peds.2009-2175
|
| [11] |
Laillou A, Prak S, De Groot R, et al. (2014) Optimal screening of children with acute malnutrition requires a change in current WHO guidelines as MUAC and WHZ identify different patient groups. PLoS One 9: e101159. doi: 10.1371/journal.pone.0101159
|
| [12] | World Health Organization (2009) WHO AnthroPlus for Personal Computers Manual Software for assessing growth of the world's children. 1-45. Available from: https://www.who.int/childgrowth/software/en/. |
| [13] | Shekhar S, Shah D (2012) Validation of mid upper arm circumference cut offs to diagnose severe wasting in Indian children. Indian Pediatr 49: 496-497. |
| [14] |
Aguayo V, Badgaiyan N, Qadir S, et al. (2018) Community management of acute malnutrition (CMAM) programme in Pakistan effectively treats children with uncomplicated severe wasting. Matern Child Nutr 14: e12623. doi: 10.1111/mcn.12623
|
| [15] |
Myatt M, Khara T, Dolan C, et al. (2019) Improving screening for malnourished children at high risk of death: A study of children aged 6–59 months in rural Senegal. Public Health Nutr 22: 862-871. doi: 10.1017/S136898001800318X
|
| [16] |
Chomtho S, Fewtrell M, Jaffe A, et al. (2006) Evaluation of arm anthropometry for assessing pediatric body composition: Evidence from healthy and sick children. Pediatr Res 59: 860-865. doi: 10.1203/01.pdr.0000219395.83159.91
|
| [17] |
Grellety E, Golden M (2016) Weight-for-height and mid-upper-arm circumference should be used independently to diagnose acute malnutrition: policy implications. BMC Nutr 2: 10. doi: 10.1186/s40795-016-0049-7
|
| [18] |
Berkley J, Mwangi I, Griffiths K, et al. (2005) Assessment of severe malnutrition among hospitalized children in rural Kenya: Comparison of Weight for Height and Mid Upper Arm Circumference. Am Med Assoc 294: 591-597. doi: 10.1001/jama.294.5.591
|
| [19] |
Myatt M, Duffield A, Seal A, et al. (2009) The effect of body shape on weight-for-height and mid-upper arm circumference based case definitions of acute malnutrition in Ethiopian children. Ann Hum Biol 36: 5-20. doi: 10.1080/03014460802471205
|
| [20] |
Katzmarzyk P, Leonard W (1998) Climatic influences on human body size and proportions: ecological adaptations and secular trends. Am J Phys Anthropol 106: 483-503. doi: 10.1002/(SICI)1096-8644(199808)106:4<483::AID-AJPA4>3.0.CO;2-K
|
| [21] | Fahmida Umiand Drupa Dillon (2011) Nutritional Assessment The Ministry of Education and Culture, the Republic of Indonesia. |
 publichealth-07-01-016-s001.pdf publichealth-07-01-016-s001.pdf |
 |
Figures(1) / Tables(1)

Tran Thi Hai, Saptawati Bardosono, Luh Ade Ari Wiradnyani, Le Thi Hop, Hoang T. Duc Ngan, Huynh Nam Phuong. The optimal mid-upper-arm circumference cutoffs to screen severe acute malnutrition in Vietnamese children[J]. AIMS Public Health, 2020, 7(1): 188-196. doi: 10.3934/publichealth.2020016







 DownLoad:
DownLoad: