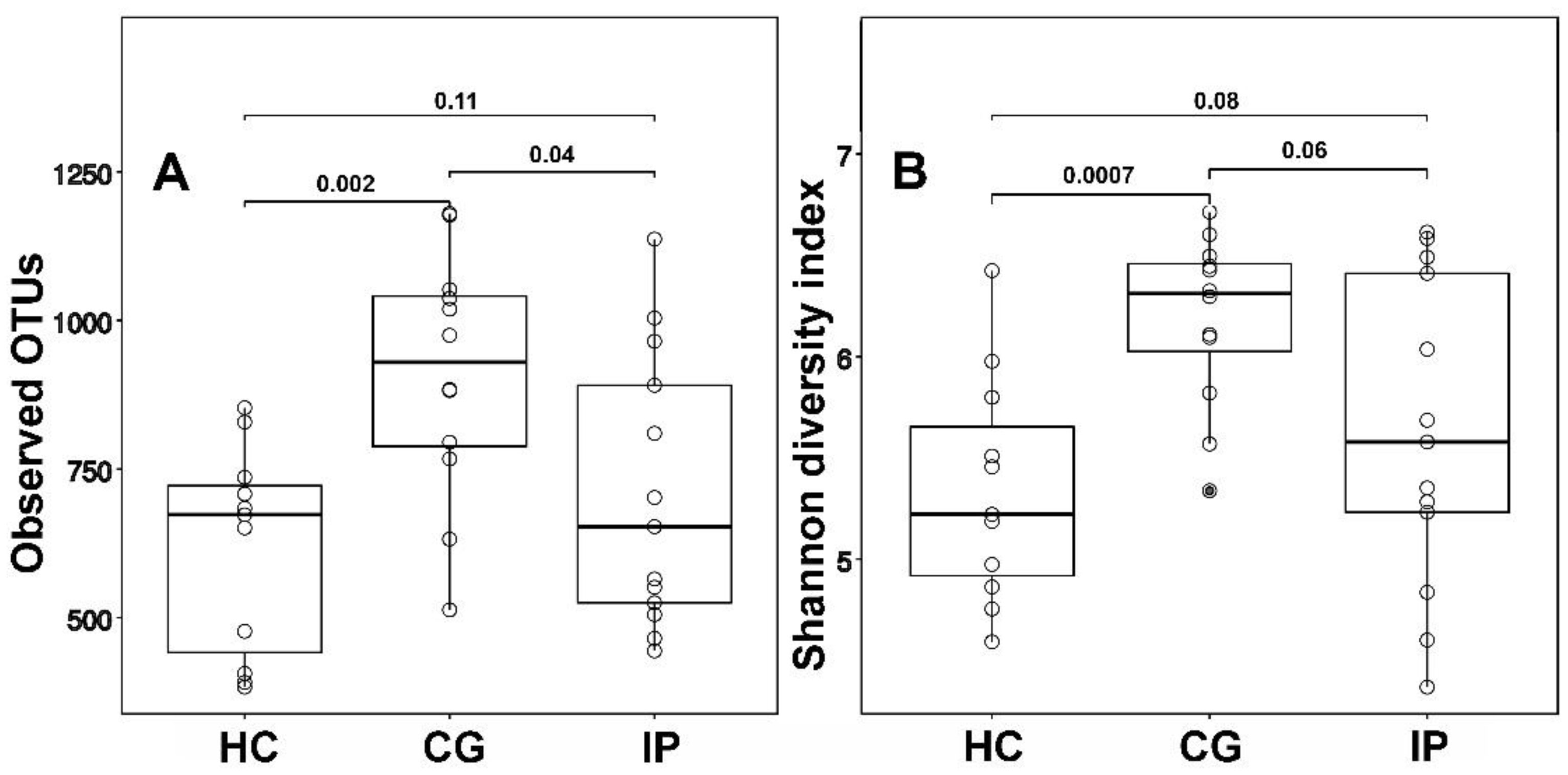
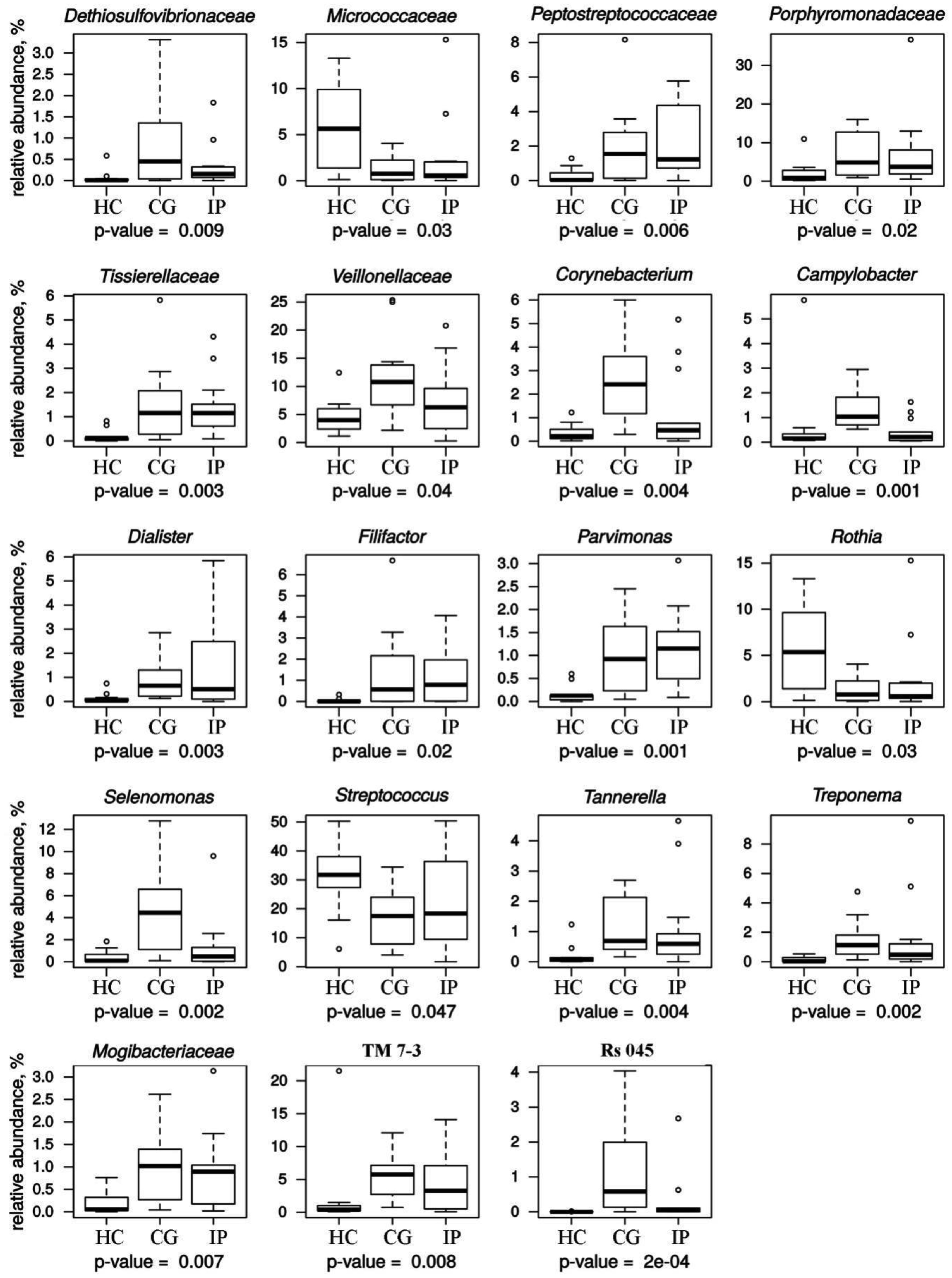
Host susceptibility and environmental factors are important for the development of gingivitis and periodontitis, but bacterial biofilms attached to the teeth and gingival tissues play a crucial role. We have analyzed and compared the subgingival microbial communities between subjects with dental plaque biofilm-induced generalized chronic gingivitis (CG), localized initial (Stage I) periodontitis (IP) and healthy controls (HC) of young people aged 18–19 years permanently residing in the city of Kazan (Tatarstan, Russia). The results showed that the α-diversity in groups with CG and IP was higher than in the healthy group. In a course of periodontal disease, a decrease in the relative abundance of dominates genera Rothia and Streptococcus was observed along with increase of class TM7-3 (Candidatus Saccharibacteria phylum) representatives. Also, the increase of red complex representatives Porphyromonadeceae, Treponema and Tannerella was detected together with statistically significant increase of Filifactor, Parvimonas, Peptostreptococcaceae, Veillonellaceae, Tissierelaceae and Mogibacteriaceae. Analysis of our data suggests that transition from HC to IP may be accompanied by a decrease in microbial diversity and a reduction in the abundance of family Rs-045 (Candidatus Saccharibacteria phylum), Desulfovibrionaceae Corynebacterium, Campylobacter and Selenomonas in young adults Kazan Tatars.
Citation: Maya Kharitonova, Peter Vankov, Airat Abdrakhmanov, Elena Mamaeva, Galina Yakovleva, Olga Ilinskaya. The composition of microbial communities in inflammatory periodontal diseases in young adults Tatars[J]. AIMS Microbiology, 2021, 7(1): 59-74. doi: 10.3934/microbiol.2021005
Host susceptibility and environmental factors are important for the development of gingivitis and periodontitis, but bacterial biofilms attached to the teeth and gingival tissues play a crucial role. We have analyzed and compared the subgingival microbial communities between subjects with dental plaque biofilm-induced generalized chronic gingivitis (CG), localized initial (Stage I) periodontitis (IP) and healthy controls (HC) of young people aged 18–19 years permanently residing in the city of Kazan (Tatarstan, Russia). The results showed that the α-diversity in groups with CG and IP was higher than in the healthy group. In a course of periodontal disease, a decrease in the relative abundance of dominates genera Rothia and Streptococcus was observed along with increase of class TM7-3 (Candidatus Saccharibacteria phylum) representatives. Also, the increase of red complex representatives Porphyromonadeceae, Treponema and Tannerella was detected together with statistically significant increase of Filifactor, Parvimonas, Peptostreptococcaceae, Veillonellaceae, Tissierelaceae and Mogibacteriaceae. Analysis of our data suggests that transition from HC to IP may be accompanied by a decrease in microbial diversity and a reduction in the abundance of family Rs-045 (Candidatus Saccharibacteria phylum), Desulfovibrionaceae Corynebacterium, Campylobacter and Selenomonas in young adults Kazan Tatars.

| [1] | Research, Science and Therapy Committee Guidelines of the American Academy of Periodontology (2001) Treatment of plaque-induced gingivitis, chronic periodontitis, and other clinical conditions. J Periodontol 72: 1790-1800. |
| [2] |
Wade WG (2013) The oral microbiome in health and disease. Pharmacol Res 69: 137-143. doi: 10.1016/j.phrs.2012.11.006

|
| [3] |
Aruni AW, Mishra A, Dou Y, et al. (2015) Filifactor alocis–a new emerging periodontal pathogen. Microbes Infect 17: 517-530. doi: 10.1016/j.micinf.2015.03.011
|
| [4] |
Kinane DF, Stathopoulou PG, Papapanou PN (2017) Periodontal diseases. Nat Rev Dis Primers 3: 17038. doi: 10.1038/nrdp.2017.38
|
| [5] |
Dewhirst FE, Chen T, Izard J, et al. (2010) The human oral microbiome. J Bacteriol 192: 5002-5017. doi: 10.1128/JB.00542-10
|
| [6] | Do T, Devine D, Marsh PD (2013) Oral biofilms: molecular analysis, challenges, and future prospects in dental diagnostics. Clin Cosmet Invest Dent 5: 11-19. |
| [7] |
Griffen AL, Beall CJ, Campbell JH, et al. (2012) Distinct and complex bacterial profiles in human periodontitis and health revealed by 16S pyrosequencing. ISME J 6: 1176-1185. doi: 10.1038/ismej.2011.191
|
| [8] |
Belstrøm D, Constancias F, Liu Y, et al. (2017) Metagenomic and metatranscriptomic analysis of saliva reveals disease-associated microbiota in patients with periodontitis and dental caries. NPJ Biofilms Microbiomes 3: 23. doi: 10.1038/s41522-017-0031-4
|
| [9] |
Hiranmayi KV, Sirisha K, Ramoji Rao MV, et al. (2017) Novel Pathogens in Periodontal Microbiology. J Pharm Bioallied Sci 9: 155-163. doi: 10.4103/jpbs.JPBS_288_16
|
| [10] | Guthmiller JM, Novak KF (2002) Periodontal Diseases. Polymicrobial Diseases Washington: ASM Press. |
| [11] |
Kriebel K, Hieke C, Müller-Hilke B, et al. (2018) Oral biofilms from symbiotic to pathogenic interactions and associated disease–connection of periodontitis and rheumatic arthritis by peptidylarginine deiminase. Front Microbiol 9: 53. doi: 10.3389/fmicb.2018.00053
|
| [12] |
Corrêa JD, Fernandes GR, Calderaro DC, et al. (2019) Oral microbial dysbiosis linked to worsened periodontal condition in rheumatoid arthritis patients. Sci Rep 9: 8379. doi: 10.1038/s41598-019-44674-6
|
| [13] |
Isola G, Matarese G, Ramaglia L, et al. (2020) Association between periodontitis and glycosylated haemoglobin before diabetes onset: a cross-sectional study. Clin Oral Invest 24: 2799-2808. doi: 10.1007/s00784-019-03143-0
|
| [14] |
Caton J, Armitage G, Berglundh T, et al. (2018) A new classification scheme for periodontal and peri-implant diseases and conditions–Introduction and key changes from the 1999 classification. J Periodontol 89: S1-S8. doi: 10.1002/JPER.18-0157
|
| [15] |
Jervøe-Storm PM, Alahdab H, Koltzscher M, et al. (2007) Comparison of curet and paper point sampling of subgingival bacteria as analyzed by real–time polymerase chain reaction. J Periodontol 78: 909-917. doi: 10.1902/jop.2007.060218
|
| [16] |
Mark Welch JL, Rossetti BJ, Rieken CW, et al. (2016) Biogeography of a human oral microbiome at the micron scale. Proc Natl Acad Sci USA 113: e791-e800. doi: 10.1073/pnas.1522149113
|
| [17] |
Suzuki N, Yoneda M, Hirofuji T (2013) Mixed Red-Complex Bacterial Infection in Periodontitis. Int J Dent 2013: 587279. doi: 10.1155/2013/587279
|
| [18] |
Settem RP, El-Hassan AT, Honma K, et al. (2012) Fusobacterium nucleatum and Tannerella forsythia induce synergistic alveolar bone less in a mouse periodontitis model. Infect Immun 80: 2436-2443. doi: 10.1128/IAI.06276-11
|
| [19] |
Kistler JO, Booth V, Bradshaw DJ, et al. (2013) Bacterial community development in experimental gingivitis. PLoS One 8: e71227. doi: 10.1371/journal.pone.0071227
|
| [20] | Balan P, Brandt BW, Chong, YS, et al. (2020) Subgingival microbiota during healthy pregnancy and pregnancy gingivitis. JDR Clin Transl Res . |
| [21] |
Kirst ME, Li EC, Alfant B, et al. (2015) Dysbiosis and alterations in predicted functions of the subgingival microbiome in chronic periodontitis. Appl Environ Microbiol 81: 783-793. doi: 10.1128/AEM.02712-14
|
| [22] |
Socransky SS, Haffajee AD, Smith C, et al. (1991) Relation of counts of microbial species to clinical status at the sampled site. J Clin Periodontol 18: 766-775. doi: 10.1111/j.1600-051X.1991.tb00070.x
|
| [23] |
Zambon JJ, Christersson LA, Slots J (1983) Actinobacillus actinomycetemcomitans in human periodontal disease. Prevalence in patient groups and distribution of biotypes and serotypes within families. J Periodontol 54: 707-711. doi: 10.1902/jop.1983.54.12.707
|
| [24] |
Dani S, Prabhu A, Chaitra KR, et al. (2016) Assessment of Streptococcus mutans in healthy versus gingivitis and chronic periodontitis: A clinic-microbiological study. Contem Clin Dent 7: 529-534. doi: 10.4103/0976-237X.194114
|
| [25] |
Larsen T, Fiehn NE (2017) Dental biofilm infections–an update. APMIS 125: 376-384. doi: 10.1111/apm.12688
|
| [26] |
Carrouel F, Viennot S, Santamaria J, et al. (2016) Quantitative molecular detection of 19 major pathogens in the interdental biofilm of periodontally healthy young adults. Front Microbiol 7: 840. doi: 10.3389/fmicb.2016.00840
|
| [27] |
Bourgeois D, David A, Inquimbert C, et al. (2017) Quantification of carious pathogens in the interdental microbiota of young caries-free adults. PLoS One 12: e0185804. doi: 10.1371/journal.pone.0185804
|
| [28] |
Camelo-Castillo AJ, Mira A, Pico A, et al. (2015) Subgingival microbiota in health compared to periodontitis and the influence of smoking. Front Microbiol 6: 119. doi: 10.3389/fmicb.2015.00119
|
| [29] | Signat B, Roques C, Poulet P, et al. (2011) Fusobacterium nucleatum in periodontal health and disease. Curr Issues Mol Biol 13: 25-36. |
| [30] |
Oscarsson J, DiRienzo J, Johansson A (2020) Editorial comments to the special issue: ‘Aggregaterbacter actinomycetemcomitans–gram-negative bacterial pathogen’. Pathogens 9: 441. doi: 10.3390/pathogens9060441
|
| [31] |
Lee WH, Chen HM, Yang SF, et al. (2017) Bacterial alterations in salivary microbiota and their association in oral cancer. Sci Rep 7: 16540. doi: 10.1038/s41598-017-16418-x
|
| [32] |
Coretti L, Cuomo M, Florio E, et al. (2017) Subgingival dysbiosis in smoker and non-smoker patients with chronic periodontitis. Mol Med Rep 15: 2007-2014. doi: 10.3892/mmr.2017.6269
|
| [33] |
Yang I, Knight AK, Dunlop AL, et al. (2019) Characterizing the subgingival microbiome of pregnant African American women. J Obstet Gynecol Neonatal Nurs 48: 140-152. doi: 10.1016/j.jogn.2018.12.003
|
| [34] |
Faveri M, Mayer MP, Feres M, et al. (2008) Microbiological diversity of generalized aggressive periodontitis by 16S rRNA clonal analysis. Oral Microbiol Immun 23: 112-118. doi: 10.1111/j.1399-302X.2007.00397.x
|
| [35] |
Brinig MM, Lepp PW, Ouverney CC, et al. (2003) Prevalence of bacteria of division TM7 in human subgingival plaque and their association with disease. Appl Environ Microbiol 69: 1687-1694. doi: 10.1128/AEM.69.3.1687-1694.2003
|
| [36] |
Islam MM, Ekuni D, Toyama N, et al. (2020) Relationship of salivary microbiome with the worsening of the periodontal health status in young adults: A 3-Year cohort study. Int J Environ Res Public Health 17: 1764. doi: 10.3390/ijerph17051764
|
| [37] |
Rylev M, Bek-Thomsen M, Reinholdt J, et al. (2011) Microbiological and immunological characteristics of young Moroccan patients with aggressive periodontitis with and without detectable Aggregatibacter actinomycetemcomitans JP2 infection. Mol Oral Microbiology 26: 35-51. doi: 10.1111/j.2041-1014.2010.00593.x
|
| [38] |
He X, McLean JS, Edlund A, et al. (2015) Cultivation of a human-associated TM7 phylotype reveals a reduced genome and epibiotic parasitic lifestyle. Proc Natl Acad Sci USA 112: 244-249. doi: 10.1073/pnas.1419038112
|
| [39] |
Bor B, Bedree JK, Shi W, et al. (2019) Saccharibacteria (TM7) in the Human Oral Microbiome. J Dent Res 98: 500-509. doi: 10.1177/0022034519831671
|
| [40] |
Soro V, Dutton LC, Sprague SV, et al. (2014) Axenic culture of a Candidate division TM7 bacterium from the human oral cavity and biofilm interactions with other oral bacteria. Appl Environ Microbiol 80: 6480-6489. doi: 10.1128/AEM.01827-14
|
| [41] |
Bedree JK, Bor B, Cen L, et al. (2018) Quorum sensing modulates the epibiotic-parasitic relationship between Actinomyces odontolyticus and its saccharibacteria epibiont, a Nanosynbacter lyticus Strain, TM7x. Front Microbiol 9: 2049. doi: 10.3389/fmicb.2018.02049
|
| [42] |
Albertsen M, Hugenholtz P, Skarshewski A, et al. (2013) Genome sequences of rare, uncultured bacteria obtained by differential coverage binning of multiple metagenomes. Nat Biotechnol 31: 533-538. doi: 10.1038/nbt.2579
|
| [43] |
Podar M, Abulencia CB, Walcher M, et al. (2007) Targeted access to the genomes of low-abundance organisms in complex microbial communities. Appl Environ Microbiol 73: 3205-3214. doi: 10.1128/AEM.02985-06
|
| [44] |
Tanner A, Maiden MF, Macuch PJ, et al. (1998) Microbiota of health, gingivitis, and initial periodontitis. J Clin Periodontol 25: 85-98. doi: 10.1111/j.1600-051X.1998.tb02414.x
|
| [45] |
Premaraj TS, Vella R, Chung J, et al. (2020) Ethnic variation of oral microbiota in children. Sci Rep 10: 14788. doi: 10.1038/s41598-020-71422-y
|
| [46] |
Mason MR, Nagaraja HN, Camerlengo T, et al. (2013) Deep sequencing identifies ethnicity-specific bacterial signatures in the oral microbiome. PLoS One 8: e77287. doi: 10.1371/journal.pone.0077287
|
| [47] |
Li J, Quinque D, Horz HP, et al. (2014) Comparative analysis of the human saliva microbiome from different climate zones: Alaska, Germany, and Africa. BMC Microbiol 14: 316. doi: 10.1186/s12866-014-0316-1
|
| [48] |
Gao L, Xu T, Huang G, et al. (2018) Oral microbiomes: more and more importance in oral cavity and whole body. Protein Cell 9: 488-500. doi: 10.1007/s13238-018-0548-1
|
| [49] |
Bourgeois D, Inquimbert C, Ottolenghi L, et al. (2019) Periodontal pathogens as risk factors of cardiovascular diseases, diabetes, rheumatoid arthritis, cancer, and chronic obstructive pulmonary disease-is there cause for consideration? Microorganisms 7: 424. doi: 10.3390/microorganisms7100424
|
| [50] |
Fan X, Alekseyenko AV, Wu J, et al. (2018) Human oral microbiome and prospective risk for pancreatic cancer: a population-based nested case-control study. Gut 67: 120-127. doi: 10.1136/gutjnl-2016-312580
|
| [51] |
Willis JR, Gabaldón T (2020) The Human Oral Microbiome in Health and Disease: From Sequences to Ecosystems. Microorganisms 8: 308. doi: 10.3390/microorganisms8020308
|
| [52] |
McHugh J (2017) Rheumatoid arthritis: New model linking periodontitis and RA. Nat Rev Rheumatol 13: 66. doi: 10.1038/nrrheum.2016.221
|
| [53] |
Stein SP, Steffen MJ, Smith C, et al. (2012) Serum antibodies to periodontal pathogens are a risk factor for Alzheimer's disease. Alzheimers Dement 8: 196-203. doi: 10.1016/j.jalz.2011.04.006
|
Figures(2) / Tables(2)

Maya Kharitonova, Peter Vankov, Airat Abdrakhmanov, Elena Mamaeva, Galina Yakovleva, Olga Ilinskaya. The composition of microbial communities in inflammatory periodontal diseases in young adults Tatars[J]. AIMS Microbiology, 2021, 7(1): 59-74. doi: 10.3934/microbiol.2021005







 DownLoad:
DownLoad: