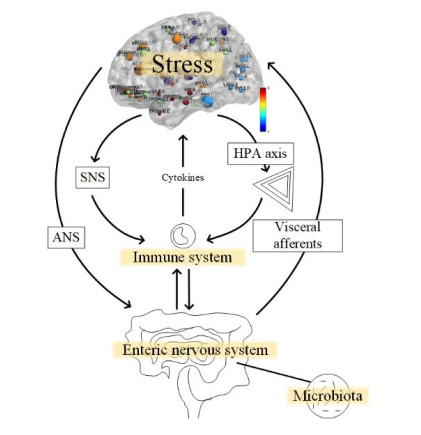
Research on functional changes in the brain of inflammatory bowel disease (IBD) patients is emerging around the world, which brings new perspectives to medical research. In this paper, the methods of canonical correlation analysis (CCA), kernel canonical correlation analysis (KCCA), and sparsity preserving canonical correlation analysis (SPCCA) were applied to the fusion of simultaneous EEG-fMRI data from 25 IBD patients and 15 healthy individuals. The CCA, KCCA and SPCCA fusion methods were used for data processing to compare the results obtained by the three methods. The results clearly show that there is a significant difference in the activation intensity between IBD and healthy control (HC), not only in the frontal lobe (p < 0.01) and temporal lobe (p < 0.01) regions, but also in the posterior cingulate gyrus (p < 0.01), gyrus rectus (p < 0.01), and amygdala (p < 0.01) regions, which are usually neglected. The mean difference in the SPCCA activation intensity was 60.1. However, the mean difference in activation intensity was only 36.9 and 49.8 by using CCA and KCCA. In addition, the correlation of the relevant components selected during the SPCCA calculation was high, with correlation components of up to 0.955; alternatively, the correlations obtained from CCA and KCCA calculations were only 0.917 and 0.926, respectively. It can be seen that SPCCA is indeed superior to CCA and KCCA in processing high-dimensional multimodal data. This work reveals the process of analyzing the brain activation state in IBD disease, provides a further perspective for the study of brain function, and opens up a new avenue for studying the SPCCA method and the change in the intensity of brain activation in IBD disease.
Citation: Yin Zhang, Xintong Wu, Jingwen Sun, Kecen Yue, Shuangshuang Lu, Bingjian Wang, Wenjia Liu, Haifeng Shi, Ling Zou. Exploring changes in brain function in IBD patients using SPCCA: a study of simultaneous EEG-fMRI[J]. Mathematical Biosciences and Engineering, 2024, 21(2): 2646-2670. doi: 10.3934/mbe.2024117
Research on functional changes in the brain of inflammatory bowel disease (IBD) patients is emerging around the world, which brings new perspectives to medical research. In this paper, the methods of canonical correlation analysis (CCA), kernel canonical correlation analysis (KCCA), and sparsity preserving canonical correlation analysis (SPCCA) were applied to the fusion of simultaneous EEG-fMRI data from 25 IBD patients and 15 healthy individuals. The CCA, KCCA and SPCCA fusion methods were used for data processing to compare the results obtained by the three methods. The results clearly show that there is a significant difference in the activation intensity between IBD and healthy control (HC), not only in the frontal lobe (p < 0.01) and temporal lobe (p < 0.01) regions, but also in the posterior cingulate gyrus (p < 0.01), gyrus rectus (p < 0.01), and amygdala (p < 0.01) regions, which are usually neglected. The mean difference in the SPCCA activation intensity was 60.1. However, the mean difference in activation intensity was only 36.9 and 49.8 by using CCA and KCCA. In addition, the correlation of the relevant components selected during the SPCCA calculation was high, with correlation components of up to 0.955; alternatively, the correlations obtained from CCA and KCCA calculations were only 0.917 and 0.926, respectively. It can be seen that SPCCA is indeed superior to CCA and KCCA in processing high-dimensional multimodal data. This work reveals the process of analyzing the brain activation state in IBD disease, provides a further perspective for the study of brain function, and opens up a new avenue for studying the SPCCA method and the change in the intensity of brain activation in IBD disease.

| [1] |
J. Kornelsen, K. Witges, J. Labus, E. A. Mayer, C. N. Bernstein, Brain structure and function changes in inflammatory bowel disease, Neuroimage Rep., 2 (2022), 100097. https://doi.org/10.1016/j.ynirp.2022.100097 doi: 10.1016/j.ynirp.2022.100097

|
| [2] |
D. C. Baumgart, W. J. Sandborn, Crohn's disease, Lancet, 380 (2012), 1590–1605. https://doi.org/10.1016/S0140-6736(12)60026-9 doi: 10.1016/S0140-6736(12)60026-9
|
| [3] |
R. J. Porter, R. Kalla, G. T. Ho, Ulcerative colitis: Recent advances in the understanding of disease pathogenesis. F1000Research, 9 (2020). https://doi.org/10.12688/f1000research.20805.1 doi: 10.12688/f1000research.20805.1
|
| [4] |
J. R. Walker, J. P. Ediger, L. A. Graff, J. M. M. A. Greenfeld, I. Clara, L. Lix, et al., The Manitoba IBD cohort study: a population-based study of the prevalence of lifetime and 12-month anxiety and mood disorders, Am. J. Gastroenterol., 103 (2008), 1989–1997. https://doi.org/10.1111/j.1572-0241.2008.01980.x doi: 10.1111/j.1572-0241.2008.01980.x
|
| [5] |
R. A. Marrie, R. Walld, J. M. Bolton, J. Sareen, J. R. Walker, S. B. Patten, et al., Increased incidence of psychiatric disorders in immune-mediated inflammatory disease, J. Psychosom. Res., 101 (2017), 17–23. https://doi.org/10.1016/j.jpsychores.2017.07.015 doi: 10.1016/j.jpsychores.2017.07.015
|
| [6] |
C. E. Whitehouse, J. D. Fisk, C. N. Bernstein, L. I. Berrigan, J. M. Bolton, L. A. Graff, et al., Comorbid anxiety, depression, and cognition in MS and other immune-mediated disorders, Neurology, 92 (2019), e406–e417. https://doi.org/10.1212/WNL.0000000000006854 doi: 10.1212/WNL.0000000000006854
|
| [7] |
L. M. Kurina, M. J. Goldacre, D. Yeates, L. E. Gill, Depression and anxiety in people with inflammatory bowel disease, J. Epidemiol. Commun. H., 55 (2001), 716–720. https://doi.org/10.1136/jech.55.10.716 doi: 10.1136/jech.55.10.716
|
| [8] |
R. Neuendorf, A. Harding, N. Stello, D. Hanes, H. Wahbeh, Depression and anxiety in patients with inflammatory bowel disease: a systematic review, J. Psychosom. Res., 87 (2016), 70–80. https://doi.org/10.1016/j.jpsychores.2016.06.001 doi: 10.1016/j.jpsychores.2016.06.001
|
| [9] |
E. Fuller-Thomson, J. Sulman, Depression and inflammatory bowel disease: findings from two nationally representative Canadian surveys, Inflamm. Bowel Dis., 12 (2006), 697–707. https://doi.org/10.1097/00054725-200608000-00005 doi: 10.1097/00054725-200608000-00005
|
| [10] |
Y. Wang, T. Chen, Q. Chen, G. Si, Emotional decisions in structured populations for the evolution of public cooperation, Physica A, 468 (2017), 475–481. https://doi.org/10.1016/j.physa.2016.10.077 doi: 10.1016/j.physa.2016.10.077
|
| [11] |
J. M. Park, J. F. Samuels, M. A. Grados, M. A. Riddle, O. Joseph Bienvenu, F. S. Goes, et al., ADHD and executive functioning deficits in OCD youths who hoard, J. Psychiatr. Res., 82 (2016), 141–148. https://doi.org/10.1016/j.jpsychires.2016.07.024 doi: 10.1016/j.jpsychires.2016.07.024
|
| [12] |
C. Tsigos, G. P. Chrousos, Hypothalamic–pituitary–adrenal axis, neuroendocrine factors and stress, J. Psychosom. Res., 53 (2002), 865–871. https://doi.org/10.1016/S0022-3999(02)00429-4 doi: 10.1016/S0022-3999(02)00429-4
|
| [13] |
E. A. Mayer, T. Savidge, R. J. Shulman, Brain–gut microbiome interactions and functional bowel disorders, Gastroenterology, 146 (2014), 1500–1512. https://doi.org/10.1053/j.gastro.2014.02.037 doi: 10.1053/j.gastro.2014.02.037
|
| [14] | J. Appleton, The gut-brain axis: Influence of microbiota on mood and mental health, Integr. Med., 17 (2018), 28–32. |
| [15] |
B. L. Bonaz, C. N. Bernstein, Brain-gut interactions in inflammatory bowel disease, Gastroenterology, 144 (2013), 36–49. https://doi.org/10.1053/j.gastro.2012.10.003 doi: 10.1053/j.gastro.2012.10.003
|
| [16] |
A. K. Thomann, M. M. Schmitgen, D. Kmuche, M. P. Ebert, P. A. Thomann, K. Szabo, et al., Exploring joint patterns of brain structure and function in inflammatory bowel diseases using multimodal data fusion, Neurogastroent. Motil., 33 (2021), e14078. https://doi.org/10.1111/nmo.14078 doi: 10.1111/nmo.14078
|
| [17] |
E. A. Mayer, Gut feelings: the emerging biology of gut–brain communication, Nat. Rev. Neurosci., 12 (2011), 453–466. https://doi.org/10.1038/nrn3071 doi: 10.1038/nrn3071
|
| [18] |
E. A. Mayer, K. Tillisch, The brain-gut axis in abdominal pain syndromes, Ann. Rev. Med., 62 (2011), 381–396. https://doi.org/10.1146/annurev-med-012309-103958 doi: 10.1146/annurev-med-012309-103958
|
| [19] |
M. H. Huang, S. Y. Fan, I. M. Lin, EEG coherences of the fronto-limbic circuit between patients with major depressive disorder and healthy controls, J. Affect. Disorders, 331 (2023), 112–120. https://doi.org/10.1016/j.jad.2023.03.055 doi: 10.1016/j.jad.2023.03.055
|
| [20] |
A. Agostini, F. Benuzzi, N. Filippini, A. Bertani, A. Scarcelli, V. Farinelli, et al., New insights into the brain involvement in patients with Crohn's disease: a voxel‐based morphometry study, Neurogastroent. Motil., 25 (2013), 147–e82. https://doi.org/10.1111/nmo.12017 doi: 10.1111/nmo.12017
|
| [21] |
A. Agostini, N. Filippini, F. Benuzzi, A. Bertani, A. Scarcelli, C. Leoni, et al., Functional magnetic resonance imaging study reveals differences in the habituation to psychological stress in patients with Crohn's disease versus healthy controls, J. Behav. Med., 36 (2013), 477–487. https://doi.org/10.1007/s10865-012-9441-1 doi: 10.1007/s10865-012-9441-1
|
| [22] |
C. Bao, P. Liu, H. Liu, X. Jin, V. D. Calhoun, L. Wu, et al., Different brain responses to electro-acupuncture and moxibustion treatment in patients with Crohn's disease, Sci. Rep., 6 (2016), 36636. https://doi.org/10.1038/srep36636 doi: 10.1038/srep36636
|
| [23] |
C. H. Bao, P. Liu, H. R. Liu, L. Y. Wu, Y. Shi, W. F. Chen, et al., Alterations in brain grey matter structures in patients with Crohn's disease and their correlation with psychological distress, J. Crohn's Colitis, 9 (2015), 532–540. https://doi.org/10.1093/ecco-jcc/jjv057 doi: 10.1093/ecco-jcc/jjv057
|
| [24] |
C. H. Bao, P. Liu, H. R. Liu, L. Y. Wu, X. M. Jin, S. Y. Wang, et al., Differences in regional homogeneity between patients with Crohn's disease with and without abdominal pain revealed by resting-state functional magnetic resonance imaging, Pain, 157 (2016), 1037. https://doi.org/10.1097/j.pain.0000000000000479 doi: 10.1097/j.pain.0000000000000479
|
| [25] |
J. Kornelsen, A. Wilson, J. S. Labus, K. Witges, E. A. Mayer, C. N. Bernstein, Brain resting-state network alterations associated with Crohn's disease. Front. Neurol., 11 (2020), 48. https://doi.org/10.3389/fneur.2020.00048 doi: 10.3389/fneur.2020.00048
|
| [26] |
K. Lv, Y. H. Fan, L. Xu, M. S. Xu, Brain changes detected by functional magnetic resonance imaging and spectroscopy in patients with Crohn's disease, World J. Gastroenterol., 23 (2017), 3607. https://doi.org/10.3748/wjg.v23.i20.3607 doi: 10.3748/wjg.v23.i20.3607
|
| [27] |
V. A. Nair, P. Beniwal-Patel, I. Mbah, B. M. Young, V. Prabhakaran, S. Saha, Structural imaging changes and behavioral correlates in patients with Crohn's disease in remission, Front. Hum. Neurosci., 10 (2016), 460. https://doi.org/10.3389/fnhum.2016.00460 doi: 10.3389/fnhum.2016.00460
|
| [28] |
A. K. Thomann, M. Griebe, P. A. Thomann, D. Hirjak, M. P. Ebert, K. Szabo, et al., Intrinsic neural network dysfunction in quiescent Crohn's Disease, Sci. Rep., 7 (2017), 11579. https://doi.org/10.1038/s41598-017-11792-y doi: 10.1038/s41598-017-11792-y
|
| [29] |
A. K. Thomann, P. A. Thomann, R. C. Wolf, D. Hirjak, C. Schmahl, M. P. Ebert, et al., Altered markers of brain development in Crohn's Disease with extraintestinal manifestations–a pilot study, PloS one, 11 (2016), e0163202. https://doi.org/10.1371/journal.pone.0163202 doi: 10.1371/journal.pone.0163202
|
| [30] |
A. W. K. Yeung, Structural and functional changes in the brain of patients with Crohn's disease: an activation likelihood estimation meta-analysis, Brain Imaging Behav., 15 (2021), 807–818. https://doi.org/10.1007/s11682-020-00291-w doi: 10.1007/s11682-020-00291-w
|
| [31] |
A. Agostini, M. Campieri, A. Bertani, A. Scarcelli, D. Ballotta, C. Calabrese, et al., Absence of change in the gray matter volume of patients with ulcerative colitis in remission: a voxel based morphometry study, BioPsychoSocial Med., 9 (2015), 1–4. https://doi.org/10.1186/s13030-014-0028-7 doi: 10.1186/s13030-014-0028-7
|
| [32] |
A. Agostini, N. Filippini, D. Cevolani, R. Agati, C. Leoni, R. Tambasco, et al., Brain functional changes in patients with ulcerative colitis: a functional magnetic resonance imaging study on emotional processing, Inflamm. Bowel Dis., 17 (2011), 1769–1777. https://doi.org/10.1002/ibd.21549 doi: 10.1002/ibd.21549
|
| [33] |
J. Kornelsen, K. Witges, J. Labus, E. A. Mayer, C. N. Bernstein, Brain structure and function changes in ulcerative colitis, Neuroimage Rep., 1 (2021), 100064. https://doi.org/10.1016/j.ynirp.2021.100064 doi: 10.1016/j.ynirp.2021.100064
|
| [34] |
J. Turkiewicz, R. R. Bhatt, H. Wang, P. Vora, B. Krause, J. S. Sauk, et al., Altered brain structural connectivity in patients with longstanding gut inflammation is correlated with psychological symptoms and disease duration, NeuroImage Clin., 30 (2021), 102613. https://doi.org/10.1016/j.nicl.2021.102613 doi: 10.1016/j.nicl.2021.102613
|
| [35] |
W. Fan, S. Zhang, J. Hu, B. Liu, L. Wen, M. Gong, et al., Aberrant brain function in active-stage ulcerative colitis patients: a resting-state functional MRI study, Front. Hum. Neurosci., 13 (2019), 107. https://doi.org/10.3389/fnhum.2019.00107 doi: 10.3389/fnhum.2019.00107
|
| [36] |
R. M. Cichy, A. Oliva, AM/EEG-fMRI fusion primer: resolving human brain responses in space and time, Neuron, 107 (2020), 772–781. https://doi.org/10.1016/j.neuron.2020.07.001 doi: 10.1016/j.neuron.2020.07.001
|
| [37] |
U. A. Kelleci, T. Calhan, A. Sahin, Z. Aydin-Ozemir, R. Kahraman, K. Ozdil, et al., Electroencephalography findings in Crohn's disease, Clin. EEG Neurosci., 50 (2019), 129–133. https://doi.org/10.1177/1550059418767589 doi: 10.1177/1550059418767589
|
| [38] |
W. Li, W. Zhang, Z. Jiang, T. Zhou, S. Xu, L. Zou, Source localization and functional network analysis in emotion cognitive reappraisal with EEG-fMRI integration, Front. Hum. Neurosci., 16 (2022), 960784. https://doi.org/10.3389/fnhum.2022.960784 doi: 10.3389/fnhum.2022.960784
|
| [39] |
T. Nguyen, T. Potter, R. Grossman, Y. Zhang, Characterization of dynamic changes of current source localization based on spatiotemporal fMRI constrained EEG source imaging, J. Neural Eng., 15 (2018), 036017. https://doi.org/10.1088/1741-2552/aa9fb2 doi: 10.1088/1741-2552/aa9fb2
|
| [40] | F. Fang, M. Houston, S. Walker, T. Nguyen, T. Potter, Y. Zhang, Underlying modulators of frontal global field potentials in emotion regulation: an EEG-informed fMRI study, in 2019 9th International IEEE/EMBS Conference on Neural Engineering (NER), (2019), 949–952. https://doi.org/10.1109/NER.2019.8716997 |
| [41] |
T. Keinänen, S. Rytky, V. Korhonen, N. Huotari, J. Nikkinen, O. Tervonen, et al., Fluctuations of the EEG‐fMRI correlation reflect intrinsic strength of functional connectivity in default mode network, J. Neurosci. Res., 96 (2018), 1689–1698. https://doi.org/10.1002/jnr.24257 doi: 10.1002/jnr.24257
|
| [42] |
A. de Cheveigné, G. M. Di Liberto, D. Arzounian, D. D. E. Wong, J. Hjortkjær, S. Fuglsang, et al., Multiway canonical correlation analysis of brain data, Neuroimage, 186 (2019), 728–740. https://doi.org/10.1016/j.neuroimage.2018.11.026 doi: 10.1016/j.neuroimage.2018.11.026
|
| [43] |
G. Lin, J. Zhang, Y. Liu, T. Gao, W. Kong, X. Lei, et al., Ballistocardiogram artifact removal in simultaneous EEG-fMRI using generative adversarial network, J. Neurosci. Meth., 371 (2022), 109498. https://doi.org/10.1016/j.jneumeth.2022.109498 doi: 10.1016/j.jneumeth.2022.109498
|
| [44] |
Z. Fang, E. Lynn, M. Huc, S. Fogel, V. J. Knott, N. Jaworska, Simultaneous EEG+ fMRI study of brain activity during an emotional Stroop task in individuals in remission from depression, Cortex, 155 (2022), 237–250. https://doi.org/10.1016/j.cortex.2022.07.010 doi: 10.1016/j.cortex.2022.07.010
|
| [45] |
Q. Guo, T. Zhou, W. Li, L. Dong, S. Wang, L. Zou, Single‐trial EEG‐informed fMRI analysis of emotional decision problems in hot executive function, Brain Behav., 7 (2017), e00728. https://doi.org/10.1002/brb3.728 doi: 10.1002/brb3.728
|
| [46] |
A. Dehghani, H. Soltanian-Zadeh, G. A. Hossein-Zadeh, Probing fMRI brain connectivity and activity changes during emotion regulation by EEG neurofeedback, Front. Hum. Neurosci., 16 (2023), 988890. https://doi.org/10.3389/fnhum.2022.988890 doi: 10.3389/fnhum.2022.988890
|
| [47] |
Y. Liu, Y. Zhang, Z. Jiang, W. Kong, L. Zou, Exploring neural mechanisms of reward processing using coupled matrix tensor factorization: A simultaneous EEG–fMRI investigation, Brain Sci., 13 (2023), 485. https://doi.org/10.3390/brainsci13030485 doi: 10.3390/brainsci13030485
|
| [48] |
X. Zhuang, Z. Yang, T. Curran, R. Byrd, R. Nandy, D. Cordes, A family of locally constrained CCA models for detecting activation patterns in fMRI, NeuroImage, 149 (2017), 63–84. https://doi.org/10.1016/j.neuroimage.2016.12.081 doi: 10.1016/j.neuroimage.2016.12.081
|
| [49] |
D. Cordes, M. Jin, T. Curran, R. Nandy, Optimizing the performance of local canonical correlation analysis in fMRI using spatial constraints, Hum. Brain Mapp., 33 (2012), 2611–2626. https://doi.org/10.1002/hbm.21388 doi: 10.1002/hbm.21388
|
| [50] |
O. Friman, J. Cedefamn, P. Lundberg, M. Borga, H. Knutsson, Detection of neural activity in functional MRI using canonical correlation analysis, Magn. Reson. Med., 45 (2001), 323–330. https://doi.org/10.1002/1522-2594(200102)45:2<323::aid-mrm1041>3.0.co;2-# doi: 10.1002/1522-2594(200102)45:2<323::aid-mrm1041>3.0.co;2-#
|
| [51] |
O. Friman, M. Borga, P. Lundberg, H. Knutsson, Adaptive analysis of fMRI data, NeuroImage, 19 (2003), 837–845. https://doi.org/10.1016/S1053-8119(03)00077-6 doi: 10.1016/S1053-8119(03)00077-6
|
| [52] |
O. Friman, M. Borga, P. Lundberg, H. Knutsson, Detection and detrending in fMRI data analysis, NeuroImage, 22 (2004), 645–655. https://doi.org/10.1016/j.neuroimage.2004.01.033 doi: 10.1016/j.neuroimage.2004.01.033
|
| [53] |
D. R. Hardoon, J. Mourao-Miranda, M. Brammer, J. Shawe-Taylor, Unsupervised analysis of fMRI data using kernel canonical correlation, NeuroImage, 37 (2007), 1250–1259. https://doi.org/10.1016/j.neuroimage.2007.06.017 doi: 10.1016/j.neuroimage.2007.06.017
|
| [54] |
V. D. Calhoun, J. Sui, Multimodal fusion of brain imaging data: a key to finding the missing link (s) in complex mental illness, Biol. Psychiat.-Cogn. N., 1 (2016), 230–244. https://doi.org/10.1016/j.bpsc.2015.12.005 doi: 10.1016/j.bpsc.2015.12.005
|
| [55] |
K. K. Lottman, D. M. White, N. V. Kraguljac, M. A. Reid, V. D. Calhoun, F. Catao, et al., Four‐way multimodal fusion of 7 T imaging data using an m CCA+ j ICA model in first‐episode schizophrenia, Hum. Brain Mapp., 39 (2018), 1475–1488. https://doi.org/10.1002/hbm.23906 doi: 10.1002/hbm.23906
|
| [56] |
I. M. Johnstone, Multivariate analysis and Jacobi ensembles: Largest eigenvalue, Tracy–Widom limits and rates of convergence, Ann. Stat., 36 (2008), 2638. https://doi.org/10.1214/08-AOS605 doi: 10.1214/08-AOS605
|
| [57] |
Z. Bao, J. Hu, G. Pan, W. Zhou, Canonical correlation coefficients of high-dimensional Gaussian vectors: Finite rank case, Ann. Stat., 47 (2019), 612–640. https://doi.org/10.1214/18-aos1704 doi: 10.1214/18-aos1704
|
| [58] | Z. Bao, J. Hu, G. Pan, W. Zhou, Canonical correlation coefficients of high-dimensional normal vectors: finite rank case, preprint, arXiv: 1407.7194. |
| [59] |
Z. Wang, L. Wang, H. Huang, Sparse additive discriminant canonical correlation analysis for multiple features fusion, Neurocomputing, 463 (2021), 185–197. https://doi.org/10.1016/j.neucom.2021.08.013 doi: 10.1016/j.neucom.2021.08.013
|
| [60] | C. Zu, D. Zhang, Sparsity preserving canonical correlation analysis, in Pattern Recognition. CCPR 2012. Communications in Computer and Information Science, Springer, (2012), 56–63. https://doi.org/10.1007/978-3-642-33506-8_8 |
| [61] |
M. J. Rosa, M. A. Mehta, E. M. Pich, C. Risterucci, F. Zelaya, A. A. T. S. Reinders, et al., Estimating multivariate similarity between neuroimaging datasets with sparse canonical correlation analysis: an application to perfusion imaging, Front. Neurosci., 9 (2015), 366. https://doi.org/10.3389/fnins.2015.00366 doi: 10.3389/fnins.2015.00366
|
| [62] |
B. Yang, J. Cao, T. Zhou, L. Dong, L. Zou, J. Xiang, Exploration of neural activity under cognitive reappraisal using simultaneous EEG-fMRI data and kernel canonical correlation analysis, Comput. Math. Methods Med., 2018 (2018). https://doi.org/10.1155/2018/3018356 doi: 10.1155/2018/3018356
|
| [63] |
J. M. Carlson, D. Foti, L. R. Mujica-Parodi, E. Harmon-Jones, G. Hajcak, Ventral striatal and medial prefrontal BOLD activation is correlated with reward-related electrocortical activity: a combined ERP and fMRI study, Neuroimage, 57 (2011), 1608–1616. https://doi.org/10.1016/j.neuroimage.2011.05.037 doi: 10.1016/j.neuroimage.2011.05.037
|
| [64] | E. Acar, Y. Levin-Schwartz, V. D. Calhoun, T. Adali, Tensor-based fusion of EEG and FMRI to understand neurological changes in schizophrenia, in 2017 IEEE International Symposium on Circuits and Systems (ISCAS), (2017), 1–4. |
| [65] |
A. Delorme, S. Makeig, EEGLAB: an open source toolbox for analysis of single-trial EEG dynamics including independent component analysis, J. Neurosci. Methods, 134 (2004), 9–21. https://doi.org/10.1016/j.jneumeth.2003.10.009 doi: 10.1016/j.jneumeth.2003.10.009
|
| [66] |
M. Amann, M. Andělová, A. Pfister, N. Mueller-Lenke, S. Traud, J. Reinhardt, et al., Subcortical brain segmentation of two dimensional T1-weighted data sets with FMRIB's Integrated Registration and Segmentation Tool (FIRST), NeuroImage-Clin., 7 (2015), 43–52. https://doi.org/10.1016/j.nicl.2014.11.010 doi: 10.1016/j.nicl.2014.11.010
|
| [67] |
G. D. Iannetti, R. K. Niazy, R. G. Wise, P. Jezzard, J. C. W. Brooks, L. Zambreanu, et al., Simultaneous recording of laser-evoked brain potentials and continuous, high-field functional magnetic resonance imaging in humans, Neuroimage, 28 (2005), 708–719. https://doi.org/10.1016/j.neuroimage.2005.06.060 doi: 10.1016/j.neuroimage.2005.06.060
|
| [68] |
S. Makeig, S. Debener, J. Onton, A. Delorme, Mining event-related brain dynamics, Trends Cogn. Sci., 8 (2004), 204–210. https://doi.org/10.1016/j.tics.2004.03.008 doi: 10.1016/j.tics.2004.03.008
|
| [69] |
R. K. Niazy, C. F. Beckmann, G. D. Iannetti, J. M. Brady, S. M. Smith, Removal of FMRI environment artifacts from EEG data using optimal basis sets, Neuroimage, 28 (2005), 720–737. https://doi.org/10.1016/j.neuroimage.2005.06.067 doi: 10.1016/j.neuroimage.2005.06.067
|
| [70] |
C. G. Yan, X. D. Wang, X. N. Zuo, Y. F. Zang, DPABI: data processing & analysis for (resting-state) brain imaging, Neuroinformatics, 14 (2016), 339–351. https://doi.org/10.1007/s12021-016-9299-4 doi: 10.1007/s12021-016-9299-4
|
| [71] |
Y. Shigemoto, H. Matsuda, K. Kamiya, N. Maikusa, Y. Nakata, K. Ito, et al., In vivo evaluation of gray and white matter volume loss in the parkinsonian variant of multiple system atrophy using SPM8 plus DARTEL for VBM, NeuroImage-Clin., 2 (2013), 491–496. https://doi.org/10.1016/j.nicl.2013.03.017 doi: 10.1016/j.nicl.2013.03.017
|
| [72] |
Z. Yang, X. Zhuang, K. Sreenivasan, V. Mishra, T. Curran, R. Byrd, et al., 3D spatially-adaptive canonical correlation analysis: Local and global methods, NeuroImage, 169 (2018), 240–255. https://doi.org/10.1016/j.neuroimage.2017.12.025 doi: 10.1016/j.neuroimage.2017.12.025
|
| [73] |
F. Divina, M. García-Torres, T. Hu, C. E. Schaerer, Soft computing for analysis of biomedical data, Comput. Math. Methods Med., 2018 (2018). https://doi.org/10.1155/2018/3902484 doi: 10.1155/2018/3902484
|
| [74] |
V. D. Calhoun, L. Wu, K. A. Kiehl, T. Eichele, G. D. Pearlson, Aberrant processing of deviant stimuli in schizophrenia revealed by fusion of fMRI and EEG data, Acta Neuropsychiatrica, 22 (2010), 127–138. https://doi.org/10.1111/j.1601-5215.2010.00467.x doi: 10.1111/j.1601-5215.2010.00467.x
|
| [75] | M. A. Akhonda, Y. Levin-Schwartz, S. Bhinge, V. D. Calhoun, T. Adali, Consecutive independence and correlation transform for multimodal fusion: Application to EEG and fMRI data, in 2018 IEEE International Conference on Acoustics, Speech and Signal Processing (ICASSP), (2018), 2311–2315. https://doi.org/10.1109/ICASSP.2018.8462031 |
| [76] |
H. L. Masson, L. Isik, Rapid processing of observed touch through social perceptual brain regions: an EEG-fMRI fusion study, J. Neurosci., 43 (2023), 7700–7711. https://doi.org/10.1523/JNEUROSCI.0995-23.2023 doi: 10.1523/JNEUROSCI.0995-23.2023
|
| [77] |
R. Mosayebi, G. A. Hossein-Zadeh, Correlated coupled matrix tensor factorization method for simultaneous EEG-fMRI data fusion, Biomed. Signal Proces., 62 (2020), 102071. https://doi.org/10.1016/j.bspc.2020.102071 doi: 10.1016/j.bspc.2020.102071
|
| [78] |
M. G. Philiastides, T. Tu, P. Sajda, Inferring macroscale brain dynamics via fusion of simultaneous EEG-fMRI, Ann. Rev. Neurosci., 44 (2021), 315–334. https://doi.org/10.1146/annurev-neuro-100220-093239 doi: 10.1146/annurev-neuro-100220-093239
|
| [79] | C. Chatzichristos, E. Kofidis, L. De Lathauwer, S. Theodoridis, S. Van Huffel, Early soft and flexible fusion of EEG and fMRI via tensor decompositions, preprint, arXiv: 2005.07134. |
| [80] |
S. M. Sadjadi, E. Ebrahimzadeh, M. Shams, M. Seraji, H. Soltanian-Zadeh, Localization of epileptic foci based on simultaneous EEG–fMRI data, Front. Neurol., 12 (2021), 645594. https://doi.org/10.3389/fneur.2021.645594 doi: 10.3389/fneur.2021.645594
|
| [81] |
X. Wang, G. Gong, N. Li, Multimodal fusion of EEG and fMRI for epilepsy detection, Int. J. Model. Simul. Sci. Comput., 9 (2018), 1850010. https://doi.org/10.1142/S1793962318500101 doi: 10.1142/S1793962318500101
|
| [82] |
A. Omidvarnia, M. A. Kowalczyk, M. Pedersen, G. D. Jackson, Towards fast and reliable simultaneous EEG-fMRI analysis of epilepsy with automatic spike detection, Clin. Neurophysiol., 130 (2019), 368–378. https://doi.org/10.1016/j.clinph.2018.11.024 doi: 10.1016/j.clinph.2018.11.024
|






Figures(7) / Tables(5)

Yin Zhang, Xintong Wu, Jingwen Sun, Kecen Yue, Shuangshuang Lu, Bingjian Wang, Wenjia Liu, Haifeng Shi, Ling Zou. Exploring changes in brain function in IBD patients using SPCCA: a study of simultaneous EEG-fMRI[J]. Mathematical Biosciences and Engineering, 2024, 21(2): 2646-2670. doi: 10.3934/mbe.2024117







 DownLoad:
DownLoad: