Citation: Yuanshi Wang, Hong Wu. Transition of interaction outcomes in a facilitation-competition system of two species[J]. Mathematical Biosciences and Engineering, 2017, 14(5&6): 1463-1475. doi: 10.3934/mbe.2017076

| [1] | [ M. Hernandez,I. Barradas, Variation in the outcome of population interactions: Bifurcations and catastrophes, J. Math. Biol., 46 (2003): 571-594. |
| [2] | [ J. Hofbauer and K. Sigmund, Evolutionary Games and Population Dynamics, Cambridge University Press, Cambridge, UK, 1998. |
| [3] | [ T. Kawai,M. Tokeshi, Variable modes of facilitation in the upper intertidal: Goose barnacles and mussels, Marine Ecology Progress Series, 272 (2004): 203-213. |
| [4] | [ T. Kawai,M. Tokeshi, Asymmetric coexistence: Bidirectional abiotic and biotic effects between goose barnacles and mussels, Journal of Animal Ecology, 75 (2006): 928-941. |
| [5] | [ T. Kawai,M. Tokeshi, Testing the facilitation-competition paradigm under the stress-gradient hypothesis: Decoupling multiple stress factors, Proceedings of the Royal Society B: Biological Sciences, 274 (2007): 2503-2508. |
| [6] | [ X. Li,H. Wang,Y. Kuang, Global analysis of a stoichiometric producer-grazer model with Holling type functional responses, J. Math. Biol., 63 (2011): 901-932. |
| [7] | [ Z. Liu,P. Magal,S. Ruan, Oscillations in age-structured models of consumer-resource mutualisms, Disc. Cont. Dyna. Systems-B, 21 (2016): 537-555. |
| [8] | [ C. Neuhauser,J. Fargione, A mutualism-parasitism continuum model and its application to plant-mycorrhizae interactions, Ecological Modelling, 177 (2004): 337-352. |
| [9] | [ S. Soliveres,C. Smit,F. T. Maestre, Moving forward on facilitation research: Response to changing environments and effects on the diversity, functioning and evolution of plant communities, Biological Reviews., 90 (2015): 297-313. |
| [10] | [ K. Tainaka, Stationary pattern of vortices or strings in biological systems: lattice version of the Lotka-Volterra model, Physical Review Letters, 63 (1989): 2688-2691. |
| [11] | [ Y. Wang,H. Wu,J. Liang, Dynamics of a lattice gas system of three species, Commun. Non. Sci. Nume. Simu., 39 (2016): 38-57. |
| [12] | [ H. Yokoi,T. Uehara,T. Kawai,Y. Tateoka,K. Tainaka, Lattice and lattice gas models for commensalism: Two shellfishes in intertidal Zone, Scientific Research Publishing, Open Journal of Ecology, 4 (2014): 671-677. |
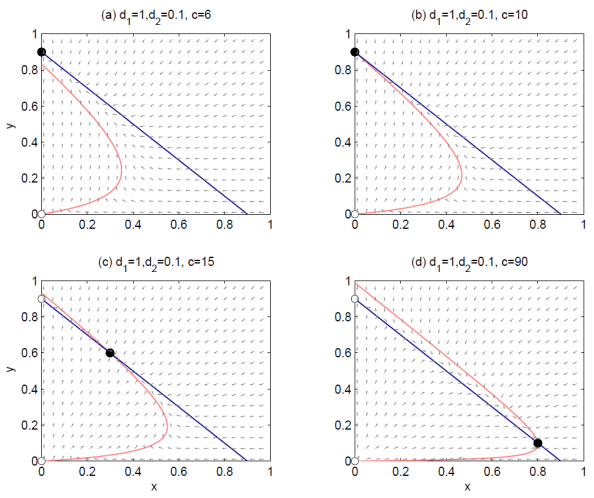
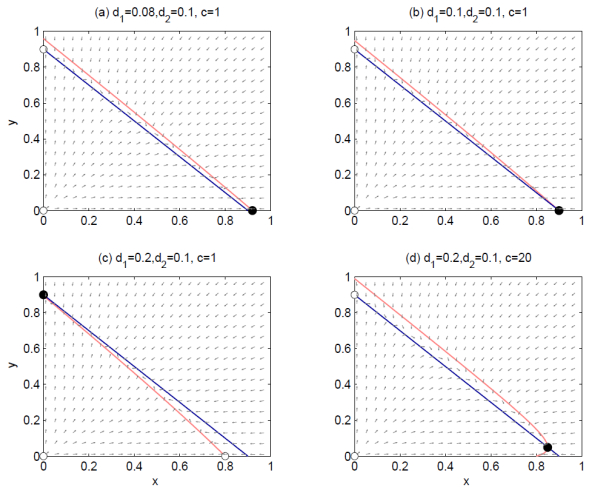
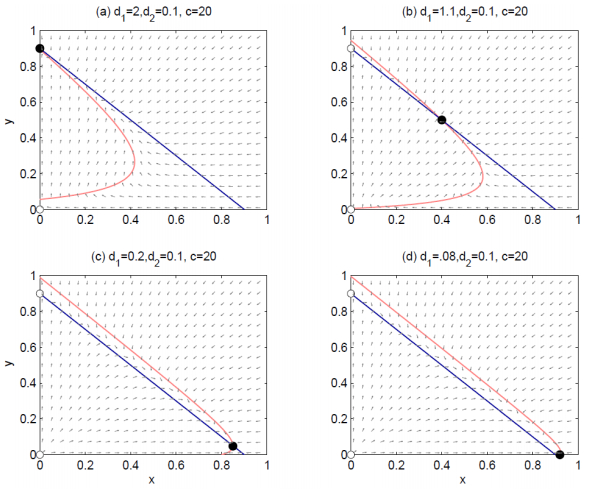
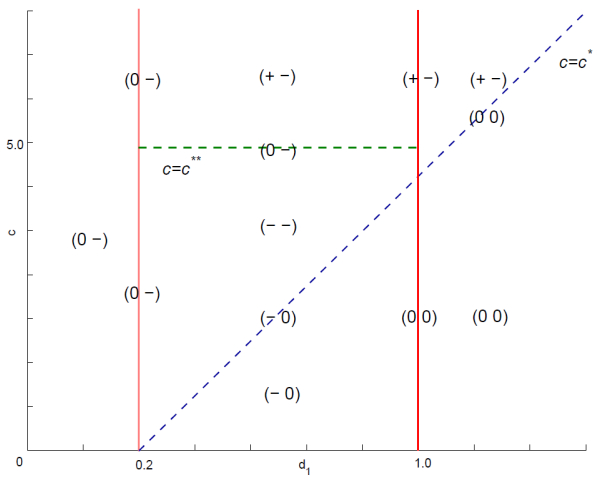
Figures(4)

Yuanshi Wang, Hong Wu. Transition of interaction outcomes in a facilitation-competition system of two species[J]. Mathematical Biosciences and Engineering, 2017, 14(5&6): 1463-1475. doi: 10.3934/mbe.2017076







 DownLoad:
DownLoad: