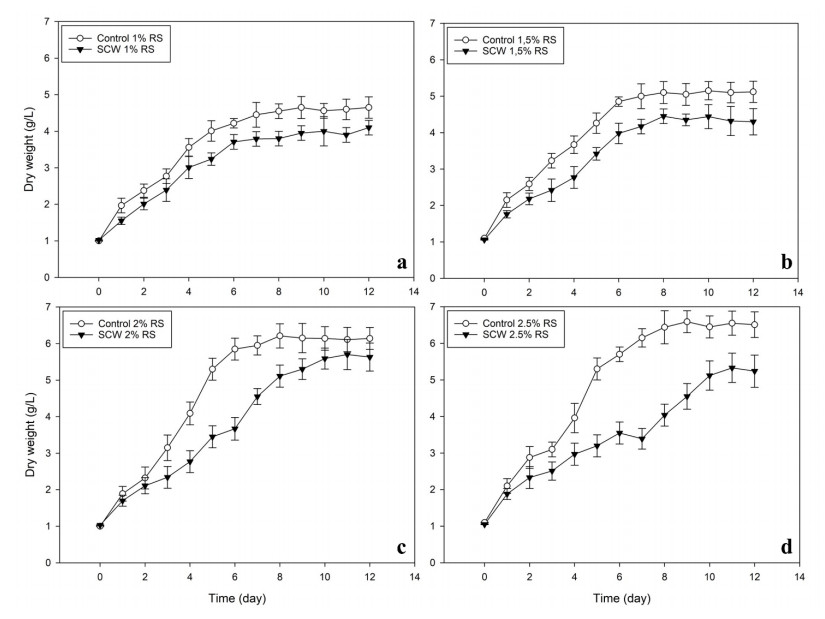
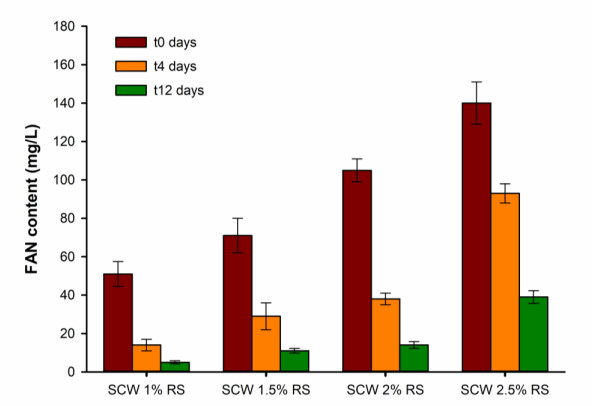
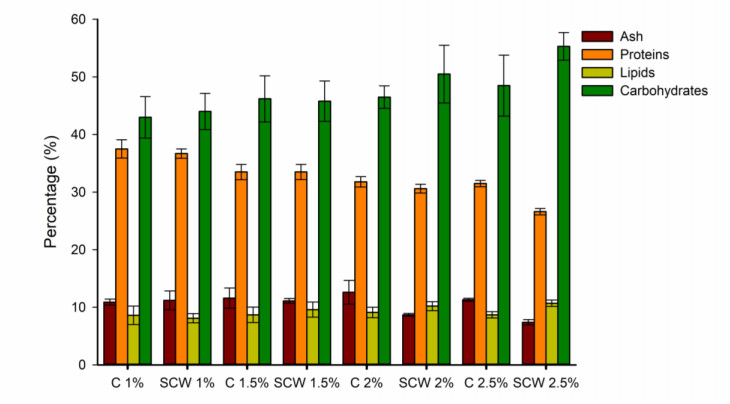
Second cheese whey (SCW) or "scotta" in Italian, is a side-stream from the manufacturing of "Ricotta" cheese, obtained after thermal coagulation of whey proteins residue in the cheese whey. Galdieria sulphuraria is a thermophilic red algae well known for its metabolic capabilities to grow on wastewater and other saline effluents. In this work, the valorisation of SCW as nutrient source for the growth of G. sulphuraria has been investigated using different concentrations of SCW. The biochemical and fatty acids composition of the biomass obtained has been evaluated too. Small differences have been observed in terms of biomass obtained after 12 days of cultivation between the SCW media and the relative control with the same amount of reducing sugars. The fatty acids composition of G. sulphuraria grown in SCW showed a higher content of polyunsaturated fatty acids compared to the control. The biomass productivity using SCW media has also been optimized through response surface methodologies with supplementation of nitrogen source obtaining a biomass dry weight higher than 10 g L-1.
Citation: Giovanni L. Russo, Antonio L. Langellotti, Maria Oliviero, Marco Baselice, Raffaele Sacchi, Paolo Masi. Valorization of second cheese whey through cultivation of extremophile microalga Galdieria sulphuraria[J]. AIMS Environmental Science, 2021, 8(5): 435-448. doi: 10.3934/environsci.2021028
Second cheese whey (SCW) or "scotta" in Italian, is a side-stream from the manufacturing of "Ricotta" cheese, obtained after thermal coagulation of whey proteins residue in the cheese whey. Galdieria sulphuraria is a thermophilic red algae well known for its metabolic capabilities to grow on wastewater and other saline effluents. In this work, the valorisation of SCW as nutrient source for the growth of G. sulphuraria has been investigated using different concentrations of SCW. The biochemical and fatty acids composition of the biomass obtained has been evaluated too. Small differences have been observed in terms of biomass obtained after 12 days of cultivation between the SCW media and the relative control with the same amount of reducing sugars. The fatty acids composition of G. sulphuraria grown in SCW showed a higher content of polyunsaturated fatty acids compared to the control. The biomass productivity using SCW media has also been optimized through response surface methodologies with supplementation of nitrogen source obtaining a biomass dry weight higher than 10 g L-1.

| [1] | Secchi N, Giunta D, Pretti L, et al. (2012) Bioconversion of ovine scotta into lactic acid with pure and mixed cultures of lactic acid bacteria. J Ind Microbiol Biotechnol. |
| [2] |
PANESAR P, KENNEDY J, GANDHI D, et al. (2007) Bioutilisation of whey for lactic acid production. Food Chem 105: 1-14. doi: 10.1016/j.foodchem.2007.03.035

|
| [3] |
Sansonetti S, Curcio S, Calabrò V, et al. (2009) Bio-ethanol production by fermentation of ricotta cheese whey as an effective alternative non-vegetable source. Biomass and Bioenergy 33: 1687-1692. doi: 10.1016/j.biombioe.2009.09.002
|
| [4] |
Tsolcha ON, Tekerlekopoulou AG, Akratos CS, et al. (2016) Treatment of second cheese whey effluents using a Choricystis -based system with simultaneous lipid production. J Chem Technol Biotechnol 91: 2349-2359. doi: 10.1002/jctb.4829
|
| [5] |
Zimermann JD ar. F, Sydney EB, Cerri ML, et al. (2020) Growth kinetics, phenolic compounds profile and pigments analysis of Galdieria sulphuraria cultivated in whey permeate in shake-flasks and stirred-tank bioreactor. J Water Process Eng 38: 101598. doi: 10.1016/j.jwpe.2020.101598
|
| [6] |
Russo GL, Langellotti AL, Oliviero M, et al. (2021) Sustainable production of food grade omega-3 oil using aquatic protists: Reliability and future horizons. N Biotechnol 62: 32-39. doi: 10.1016/j.nbt.2021.01.006
|
| [7] |
Massa M, Buono S, Langellotti AL, et al. (2019) Biochemical composition and in vitro digestibility of Galdieria sulphuraria grown on spent cherry-brine liquid. N Biotechnol 53: 9-15. doi: 10.1016/j.nbt.2019.06.003
|
| [8] |
Girard J-M, Roy M-L, Hafsa M Ben, et al. (2014) Mixotrophic cultivation of green microalgae Scenedesmus obliquus on cheese whey permeate for biodiesel production. Algal Res 5: 241-248. doi: 10.1016/j.algal.2014.03.002
|
| [9] |
Abreu AP, Fernandes B, Vicente AA, et al. (2012) Mixotrophic cultivation of Chlorella vulgaris using industrial dairy waste as organic carbon source. Bioresour Technol 118: 61-66. doi: 10.1016/j.biortech.2012.05.055
|
| [10] |
Ende SSW, Noke A (2019) Heterotrophic microalgae production on food waste and by-products. J Appl Phycol 31: 1565-1571. doi: 10.1007/s10811-018-1697-6
|
| [11] |
Scherhag P, Ackermann J (2021) Removal of sugars in wastewater from food production through heterotrophic growth of Galdieria sulphuraria. Eng Life Sci 21: 233-241. doi: 10.1002/elsc.202000075
|
| [12] |
Pan S, Dixon KL, Nawaz T, et al. (2021) Evaluation of Galdieria sulphuraria for nitrogen removal and biomass production from raw landfill leachate. Algal Res 54: 102183. doi: 10.1016/j.algal.2021.102183
|
| [13] | Moon M, Mishra SK, Kim CW, et al. (2014) Isolation and characterization of thermostable phycocyanin from Galdieria sulphuraria. Korean J Chem Eng. |
| [14] | Allen MM (1968) SIMPLE CONDITIONS FOR GROWTH OF UNICELLULAR BLUE-GREEN ALGAE ON PLATES. J Phycol 4: 1-4. |
| [15] |
Miller GL (1959) Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal Chem 31: 426-428. doi: 10.1021/ac60147a030
|
| [16] |
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248-254. doi: 10.1016/0003-2697(76)90527-3
|
| [17] | Adams VD (2017) Water and Wastewater Examination Manual. |
| [18] |
Lie S (1973) THE EBC-NINHYDRIN METHOD FOR DETERMINATION OF FREE ALPHA AMINO NITROGEN. J Inst Brew 79: 37-41. doi: 10.1002/j.2050-0416.1973.tb03495.x
|
| [19] |
Nazir Y, Shuib S, Kalil MS, et al. (2018) Optimization of Culture Conditions for Enhanced Growth, Lipid and Docosahexaenoic Acid (DHA) Production of Aurantiochytrium SW1 by Response Surface Methodology. Sci Rep 8: 8909. doi: 10.1038/s41598-018-27309-0
|
| [20] |
DuBois M, Gilles KA, Hamilton JK, et al. (1956) Colorimetric Method for Determination of Sugars and Related Substances. Anal Chem 28: 350-356. doi: 10.1021/ac60111a017
|
| [21] |
Bligh EG, Dyer WJ (1959) A RAPID METHOD OF TOTAL LIPID EXTRACTION AND PURIFICATION. Can J Biochem Physiol 37: 911-917. doi: 10.1139/o59-099
|
| [22] | AOAC (2005) Official Methods of Analysis of AOAC International. |
| [23] |
Tsolcha ON, Tekerlekopoulou AG, Akratos CS, et al. (2016) Treatment of second cheese whey effluents using a Choricystis -based system with simultaneous lipid production. J Chem Technol Biotechnol 91: 2349-2359. doi: 10.1002/jctb.4829
|
| [24] |
Oesterhelt C, Schnarrenberger C, Gross W (1999) Characterization of a sugar/polyol uptake system in the red alga Galdieria sulphuraria. Eur J Phycol 34: 271-277. doi: 10.1080/09670269910001736322
|
| [25] | Gross W, Schnarrenberger C (1995) Heterotrophic Growth of Two Strains of the Acido-Thermophilic Red Alga Galdieria sulphuraria. Plant Cell Physiol. |
| [26] |
Rigano C, Fuggi A, Rigano VDM, et al. (1976) Studies on utilization of 2-ketoglutarate, glutamate and other amino acids by the unicellular alga Cyanidium caldarium. Arch Microbiol 107: 133-138. doi: 10.1007/BF00446832
|
| [27] |
Ho S-H, Li P-J, Liu C-C, et al. (2013) Bioprocess development on microalgae-based CO2 fixation and bioethanol production using Scenedesmus obliquus CNW-N. Bioresour Technol 145: 142-149. doi: 10.1016/j.biortech.2013.02.119
|
| [28] |
Wen Z-Y, Chen F (2003) Heterotrophic production of eicosapentaenoic acid by microalgae. Biotechnol Adv 21: 273-294. doi: 10.1016/S0734-9750(03)00051-X
|
| [29] |
Tischendorf G, Oesterhelt C, Hoffmann S, et al. (2007) Ultrastructure and enzyme complement of proplastids from heterotrophically grown cells of the red alga Galdieria sulphuraria. Eur J Phycol 42: 243-251. doi: 10.1080/09670260701437642
|
| [30] |
Sloth JK, Jensen HC, Pleissner D, et al. (2017) Growth and phycocyanin synthesis in the heterotrophic microalga Galdieria sulphuraria on substrates made of food waste from restaurants and bakeries. Bioresour Technol 238: 296-305. doi: 10.1016/j.biortech.2017.04.043
|
| [31] |
Graziani G, Schiavo S, Nicolai MA, et al. (2013) Microalgae as human food: chemical and nutritional characteristics of the thermo-acidophilic microalga Galdieria sulphuraria. Food Funct 4: 144-152. doi: 10.1039/C2FO30198A
|
| [32] |
Wan M, Wang Z, Zhang Z, et al. (2016) A novel paradigm for the high-efficient production of phycocyanin from Galdieria sulphuraria. Bioresour Technol 218: 272-278. doi: 10.1016/j.biortech.2016.06.045
|
| [33] |
Sakurai T, Aoki M, Ju X, et al. (2016) Profiling of lipid and glycogen accumulations under different growth conditions in the sulfothermophilic red alga Galdieria sulphuraria. Bioresour Technol 200: 861-866. doi: 10.1016/j.biortech.2015.11.014
|
| [34] |
Martinez-Garcia M, van der Maarel MJEC (2016) Floridoside production by the red microalga Galdieria sulphuraria under different conditions of growth and osmotic stress. AMB Express 6: 71. doi: 10.1186/s13568-016-0244-6
|
Figures(3) / Tables(5)

Giovanni L. Russo, Antonio L. Langellotti, Maria Oliviero, Marco Baselice, Raffaele Sacchi, Paolo Masi. Valorization of second cheese whey through cultivation of extremophile microalga Galdieria sulphuraria[J]. AIMS Environmental Science, 2021, 8(5): 435-448. doi: 10.3934/environsci.2021028







 DownLoad:
DownLoad: